Government of the Autonomous City of Buenos Aires
Neuropsychiatric Hospital "José Tiburcio
Borda"
Laboratory of
Electroneurobiological Research
and Journal
Electroneurobiology
ISSN: 0328-0446

The
Comments on Professor Christfried Jakob's
Contributions
made in

The Cytoarchitectonics of the Adult Human
Cortex
by
Professor Constantin, Baron von Economo
in Vienna, and
Dr. Georg N. Koskinas
emeritus Assistant
of the Psychiatric and Neurological University Clinic in Athens
Created at the
Psychiatric Clinic, Director Councillor J. Wagner v. Jauregg
Vienna
Vienna and Berlin
Publisher: Julius Springer Verlag
1925
Translated into English by H. Lee Seldon (Monash Univ., Australia)
who also offers the German text of the entire book
with its illustrations, as well as his English rendering (under completion), in
his website http://neptune.netcomp.monash.edu.au/staff/lseldon/LeePublications.html
Preliminary online version (not yet completely
revised). Notes in the present article are by Mariela Szirko
Electroneurobiología
2005; 13 (1), pp. 46 - 73; URL
<http://electroneubio.secyt.gov.ar/index2.htm>
Copyright © Public.
This is a research work of public access, with redistribution granted on the
condition of conserving this notice in full and complete reference to its
publication including URLs. - Esta es una investigación de acceso público; su copia exacta y
redistribución por cualquier medio están permitidas bajo la condición de
conservar esta noticia y la referencia completa a su publicación incluyendo la
URL original (ver arriba).
Printing
this .htm file does not keep tables and original page numbers
You
can download a .PDF
(recommended: 0.87 MB) or .DOC
(1.29 MB) file for printing or better reading the footnotes, from / Acceso de
red permanente: puede obtener un archivo .PDF
(recomendado: 0,87 MB) o .DOC (1,29
MB) para leer mejor o imprimir esta investigación, de http://electroneubio.secyt.gov.ar/index2.html
Professors von Economo and Koskinas made their
commentaries to refer (Literature,
page 803) to
JAKOB, CHR.: Vom Tierhirn zum Menschenhirn, München:
Lehmann, 1911
and
Das Menschenhirn, München: Lehmann.


Christfried Jakob (1866-1956) and
Constantin Freiherr von Economo (1876-1931)
--------
General part: General basis of the cytoarchitectonics
of the cerebral cortex
Chapter 1. Introductory remarks
A. Introduction
(Page 2)
Like
the sensory organs (for example, the eye bubbles from the mesencephalon), the
hemisphere pouchs of the cerebrum develop in pairs from the single telencephalon,
and one could understand them as a sensory organ whose view is on the inner
events in the central nervous system. The stimuli which enter this organ do not
come directly from the periphery, but are merely internal stimuli that come
from the entire remaining nervous system, to be received and processed as a
total. The cerebral cortex is also capable of accumulating these stimuli, so
that the surplus part of stimulus energy, which is not used in the simple
reflex arc, collects in the brain. By being able to change past energy into
present and future energy, it frees the organism from the brutal primitive law
of the reflex act and gives him individual freedom and personality (CHR.
JAKOB).

Prof. Jakob
sectioning a human brain on the sun-drenched veranda by the South entrance of
this Laboratory (right side of the photograph), at the time (1906-7) he was
composing his interference model of macro- and microcircuits for the
installments of "Localization of the soul and intelligence". Image
added for this article
B. Historical Notes
Original pagination (from the Table of
Contents):
CHRISTFRIED JAKOB Fundamental layers, 20
His “original gyri” and sector theory, 23
Page 17
Three
names must still be mentioned, that, although they are not directly connected
with cytoarchitectonics, will still greatly influence its study, namely CAJAL,
KAES and CHRISTFRIED JAKOB.
In 1886
GOLGI gave us, with his silver impregnation method for neurons, an unique means to recognize the form of a neuron together
with its dendrites and axon. Thus, we can derive basic knowledge about the
different cells types which appear in the nervous system. Soon therefrom CAJAL
began to systematically explore the human and animal cerebral cortex by means
of various silver methods, some also wonderfully developed by him. We owe the
knowledge that we have about it today to this highly-deserving Spanish scholar.
In the discussion of the individual cells forms in Chapter 2 (cf. p. 44 - 68),
furthermore with the discussion of the individual Areas and in many other
places we will still come back to individual results of his extensive
examinations. Knowledge of the entire cortical architectonics can help us
understand the processes in it only in conjunction with the knowledge of
CAJAL's explorations of the structure of individual cells and their precise
connections projections. With regret we must register the feeling that CAJAL's
great studies were ahead of their time, as he did them before the area division
of the cortex was postulated by MEYNERT and BETZ, a postulate which would
create the necessary coarse basis for CAJAL's detailed examinations. It is
often difficult today to utilize the important results that have been provided
by silver stains, because localization of these results to precise positions in
the cortex cannot be done. Therefore, CAJAL, with his untiring creativity, has
recently started with silver impregnation of the individual "Areae"
of the cortex, and we expect extraordinarily important results from these
studies, especially for a future fibrillo-architectonics.
In 1907
KAES published a text and atlas on the normal and pathological cortex, stained
by means of the myelin method. We want to summarize the most important results
here: The cortical thickness is greater in the newborn in the first months of
life than in adults; from the third month of life to the end of the first year,
it rapidly decreases; the decrease progresses slowly and further until the end
of the 20th year of life; around the 20th year it begins to increase again and
reaches its maximum in the 5th decade of life, in order to then decrease again.
Fig. 13, curve I, p. 21, shows this behavior in excerpts from KAES' original
pictures. The gyral cap, the gyral wall and the gyral valley behave rather
uniformly here. But not all parts of the cortex participate identically in
these alterations. KAES therefore divides the cortex into a so-called outer
main layer, including the outer three MEYNERT's layers up to the outer
Baillarger stripe, and the underlying inner main layer. KAES's curve II, Fig.
13, shows that the changes of the total cortical thickness are based
specifically on fluctuations of the outer main layer, that decreases up to the
20th year and then grows significantly again up to the 45th. Actually, the
inner main layer (curve III, Fig. 13) increases progressively but very slowly
from birth up to the fifth decade of life. If this observation should prove to
be a rule, it would be a fundamental fact of the development of the brain during
life, one whose importance is immediately clear to everyone. According to KAES,
individual brain regions adhere to this curve quite differently. It is
applicable to the entire forebrain; however, it does not apply to the visual
cortex - here the development curve shows a more continuous development. In
certain brain regions, the peak of the development curve shifts to other ages;
and so each brain area apparently has its own curve. In KAES's original work
the regional alterations are caused more through the outer main layer than the
one. KAES also determined the number of projection bundles per millimeter-wide
section of the cortex at different ages. We show parts of his curves in Fig.
14. The maximum is reached at approximately the 20th year; however, again both
the number of projection bundles as well as the year of the maximum vary
regionally; the cortex of the anterior central gyrus and the visual cortex
deviate the most from this average curve. KAES thinks furthermore that the
narrower cortex is the more developed and fiber-rich; in the adult this is
usually the left side. Because of the particular development pattern and the
delayed development peak of the outer main layer (in the 5th decade of life!),
KAES believes that it plays a special role in the development of individuality
and higher intellect. One objection to the measurements of KAES is that his
numbers are too large - he gives an average of 4.9 mm for the width of the
cortex at the gyral caps of the convex surface – it should be at most 3.5 mm! -
or too inaccurate. Certainly it would be very desirable to control whether the
rules formulated by KAES retain their validity after a correction of the
measurements. Then it is certain that these rules, particularly regarding the
behavior the outer and inner main layers, would be of fundamental importance.
With the Gudden method NISSL showed experimentally that only the cells of the
inner main layer are connected to the deep ganglia and projection tracks; this
discovery also points out a fundamental difference between outer and inner main
layers. We shall see how much these layers show regional differences in Chapter
4 (cf. p. 116 - 178).

Professor von Economo, early in his career as a scientist
Page 22
(even header): Introductory remarks.
In
CHRISTFRIED JAKOB's still unfinished works "Vom Tierhirn zum
Menschenhirn" and "The Human Brain" there are quite new results.
Although like the aforementioned ones, these are not directly connected with
cytoarchitectonics, they can still influence the latter. For the outer and the
inner main layers, which he considers the two fundamental layers of the fully
developed cortex, through phylogenetic studies and examinations of Gymnophions
(Coecilis lumbricoides) - an
especially suitable object, with a brain structure between amphibians and
reptiles [Note from Jakob's Laboratory, September,
2005: a few years later, Jakob discovered an error in systematics – the
supposed gymnophions were actually amphisbaenids! Prof. Jakob treated the error
humorously throughout his life and reported it in a series of books and
letters, none of which seem to have been known to Professors von Economo and
Koskinas by the time of writing their treatise, completed in September, 1924.
As the systematic position of his observations was duly corrected, the blunder
had no neurobiological consequences. MS] - he could demonstrate a
different origin for each layer. We borrow the following explanations and
illustrations from his book. With amphibians the cerebrum comprises only the
rhinencephalon and the Striatum, and the cerebral pouch that stretches itself
over it is still purely ependymal (Fig. 15). With Coecilia [Amphisbaena;
MS], where this blanket has already developed to a wider, nervous tissue, the
neurons of this formation (Archipallium), that will become Ammon's horn in
higher animals, correspond only to the inner fundamental layer. Those at the
lateral base of the Archipallium remain in continuous contact with the cells of
the Striatum c. st. (Fig. 16a, si). However, at the place (f.m.) where the
actual rhinencephalon (Rh) is bordered in the Fissura marginalis, a lateral
cells row originating from the cells of this rhinencephalon (se) pushes itself
over the cells of the Striatum and the inner fundamental layer (si) and forms
the basis of the outer fundamental layer (se). Together they form the ordinary
cortex, the neopallium. With embryological studies of the central nervous
system of opossums, CHR. JAKOB found places which seem to support this being a
general principle (Fig. 16b). (Compare Fig. 66 image VI of the three-month
human fetus.) He infers that the outer main or fundamental layer (II + III of
MEYNERT) derives originally from the rhinencephalon and is more sensory in
nature, whereas the inner fundamental layer (V + VI), which originates from the
Striatum, is motor in nature. In later life the two unify through layer IV,
whose granular cells form a system of short associations between the two
fundamental layers. The cortex of the Archipallium, which remains relatively
constant throughout animal phylogeny, forms Ammon's horn. The lateral pouch
with the two fundamental layers becomes the neopallium (the actual gray cortex)
through strong growth dorsally and medially and through increase in width. The
always peculiarly built Insula cortex (with the Claustrum) develops from the
area of the marginal fissure. Furthermore, at the base the
"rhinencephalon" has its own further development. The neopallium
develops immensely from outside to inside and folds itself in longitudinal
pleats, the origins of gyri. The most inner one is Ammon's horn, then the Gyrus
limbicus and towards the outside - still recognizable as primitive gyri in the
dog brain - Gyrus ectomarginalis, suprasylvicus, ectosylvicus and insulae. The
Operculum is created through swelling of the cortex edge at the Fissura
marginalis. Besides this ventro-dorsal development, a fan-like unfolding of the
cortex appears in the frontocaudal direction, with a rotation point in the
Insula area. This development causes, beside the above-mentioned segmentation
in primitive gyri, a sector-shaped construction along the longitudinal axis.
This is still very clear in the cortical structure of lissencephalic animals.
Fig. 17 (CHR. JAKOB) clearly shows this. Through further fan-shaped development
posteriorly the occipital lobe arises, and through further twisting of this
rear end downward and again forward the temporal lobe arises. This is described
by the sector diagram of primates (Fig. 18, from JAKOB). Each of these sectors
has its own physiological functions and own anatomical connections. A glance at
Fig. 18 and on our brain map (Figs. 19 and 20, which we show reduced for
comparison) shows a certain astounding similarity of both. The same holds for a
comparison of Fig. 17 with the brain map of lissencephalic animals (Fig. 104,
p. 243). The future will show whether these new and basic thoughts of CHR.
JAKOB on the fundamental layers and the sector development are right. We
mentioned them extensively here, because this description of the main layers is
closely related to our architectural studies and because it is possible that
the similarity of the sector-shaped development and the borders of the Areas
that appear on these illustrations, is based on more than a mere coincidence.
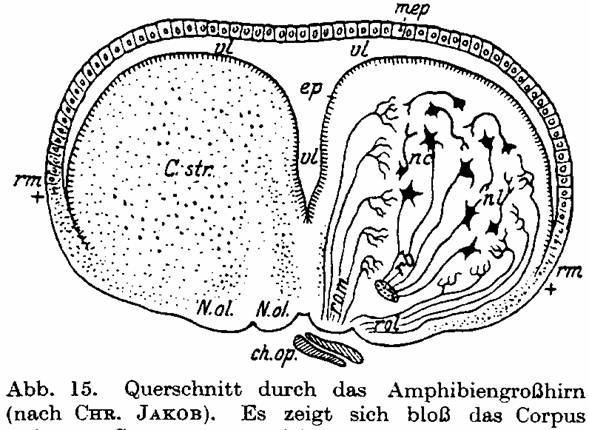
Fig. 15. Cross-section of the amphibian
cerebrum from CHR. JAKOB. The Corpus striatum C.str is well developed, while
the hemisphere cover (Pallium) appears only as a thin ependymal film mep over
the ventricle vl.
Historical Notes
(odd-page header): 23
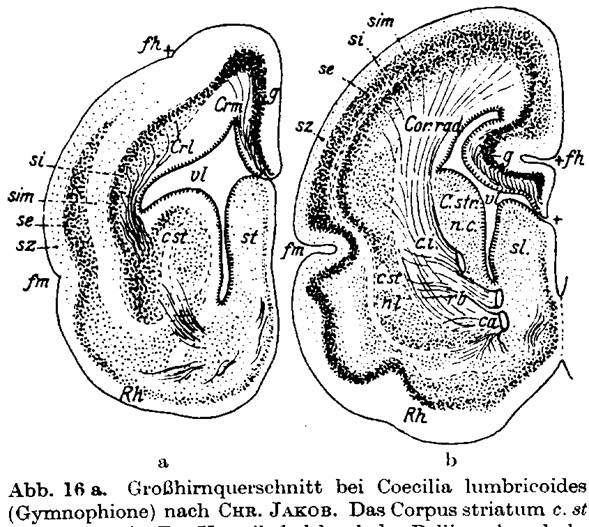
Fig. 16 a. Cerebrum cross-section of
Coecilis lumbricoides (Gymnophion) from CHR. JAKOB. [Indeed
Amphisbaenidae; see note in text. The misattributed genus' nomen is Caecilia L. 1758, from Pliny the Elder;
occasional Coecilia appears from
1790's on] The Corpus striatum c. St is well developed. The Pallium
closes the ventricle vl dorsally; it is admittedly thin, but already neurons
are present in g (Archipallium). These cells originate from the lateral band of
the Corpus striatum and form the Stratum internum si, later to become the inner
fundamental layer. Rh rhinencephalon; fm Fissura marginalis is the base of the
rhinencephalon. From here a cells row se, the later outer fundamental layer,
grows from lateral and basal dorsally over the si. Therefore, se originally
comes from the rhinencephalon and later merges with the si, which comes from
the Striatum. sz Stratum zonal, sim Stratum intermedium, fh Fissura hippocampi.
- Fig. 16b shows similar relations in a cerebrum cross-section of an embryo of
the opossum (CHR. JAKOB).
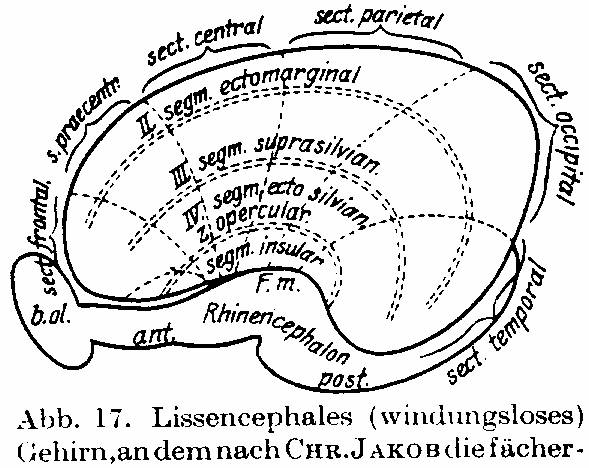
Fig. 17. Lissencephalic brain on which,
according to CHR. JAKOB, the fan-shaped development of the sectors in
frontocaudal development is drawn. The Insula forms the rotation point of this
development. Also the segmental arrangement is drawn.
24 Introductory remarks.
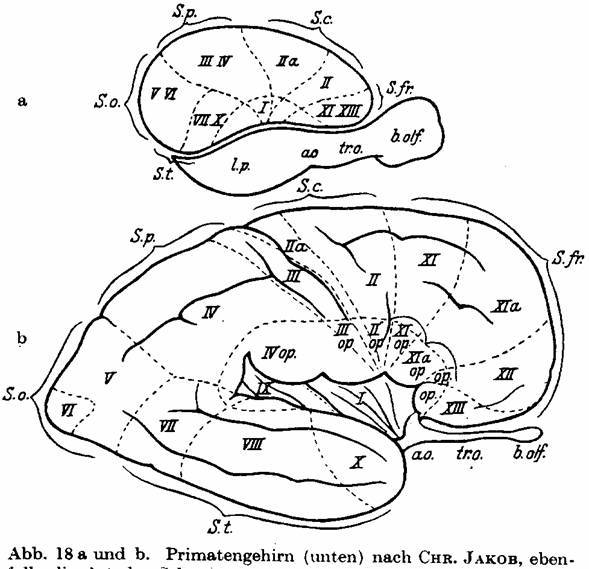
Fig. 18 a and b. Primate brain (below) from
CHR. JAKOB also shows the sector development of a sophisticated gyrencephalic
brain. The temporal lobe is pushed downward and forwards through the fan-shaped
growth, and the occipital lobe is moved to the back. - For comparison a
lissencephalic brain is shown above in order to emphasize the movement of the
sectors.
All these
studies laid the foundations of normal cortex architectonics - for the various
purposes and goals discussed above. Major among these was to create the normal
basis necessary for recognition of pathological changes, although the study of
the latter has taken place simultaneously. BETZ and HAMMARBERG already studied
brains from idiots, and KAES included such from criminals. CAMPBELL and later
SCHRÖDER made cytoarchitectonic examinations with pyramidal tract lesions and
amyotrophic lateral sclerosis. KÖLPIN and LEWY with Huntington's chorea,
SPIELMEYER and BIELSCHOWSKY with paralyses without pyramidal tract lesions,
JOSEPH, A. JAKOB, BUSCAINO and KLARFELD, DOUTREBENTE and MARCHAND with Dementia
praecox (catatonia). ALZHEIMER, BRATZ, POLLACK and KOGERER showed changes in cells
and layers with epilepsy. C. and O. VOGT have tried to create bases for a
future patho-architectonics in a detailed treatise (Illnesses of the Cerebral
Cortex), which also includes numerous good pictures of normal cortex sections.
Several detailed publications have appeared from the Viennese neurological institute of Professor MARBURG, under
his direction and with the assistance of Dr. POLLAK. They treat the
Patho-architectonics of the psychoses systematically and in detail, SAITO on
progressive paralysis, TAKASE on manic depression, NAITO on schizophrenia, and
OSAKI on senile psychoses. Earlier, WADA had studied this problem in this
institute. The first of these works is, as a basis for the judgment of
pathological changes, a rich selection of very good photographic pictures of
all BRODMANN's fields with a concise and apt description of each. So, we see
how pathology awaits with a justified and healthy impatience an exhaustive
description of the normal structures of the cortex.
Historical Notes. 25
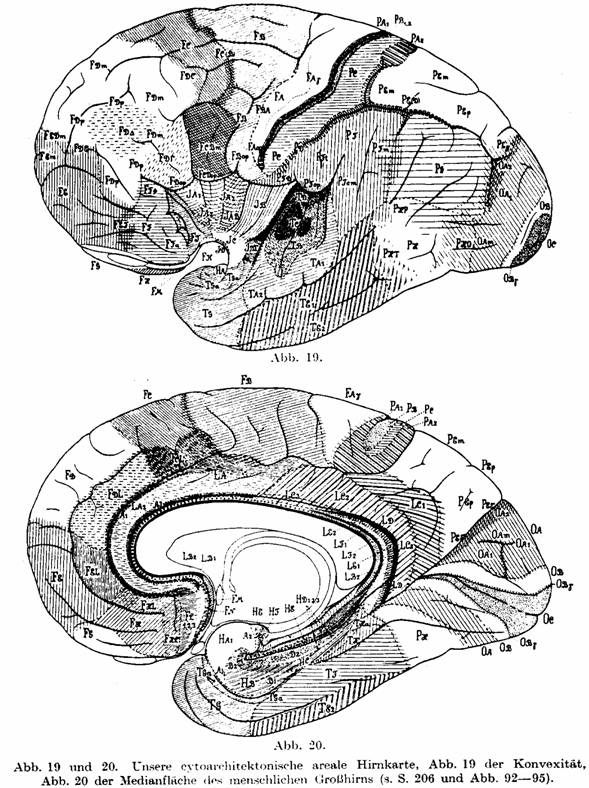
Fig. 19 and 20. Our cytoarchitectonic brain map. Fig.
19 of the convex surface, Fig. 20 of the median surface of the human cerebrum
(cf. p. 206 and Figs. 92 - 95).
Cortical
measurements Page 41
3. Cortex volumes.
The
ratio of gray cortex to white matter volume decreases with higher ranks in the
animal phylogeny. We can see this for ourselves through a glance at a brain
slice of the rabbit (Fig. 31), in which the gray cortex is extremely broad, and
the white matter mass forms only a quite small inner section (cf. Fig. 25 of
the human). But even in a comparison of a brain cross-section of a lower
monkey, then an orangutan and a human, one is able to see this progressive
increase of white matter mass and relative decrease and thinning of the cortex.
According to CHR. JAKOB, in cross-sections from lower monkeys the gray matter
prevails over the white in the ratio of 5:1; with the Orangutan only by 3:1,
and with the human approximately 2:1.
Page
42 General remarks on the cortex and its neurons.
JAEGER has
measured the volumes of the gray cortex and white matter of the hemispheres. He
determined the volume for brain slices of a certain thickness by means of
ANTON's planimetric measurements. He calculated the volume of the cortex of
both hemispheres at 540 - 580 cm3, and that of the white matter at 400 - 490
cm3 (without the medulla). On average the ratio would be 560:445 or
approximately 1.2: 1. According to DANILEWSKI, the density of the gray matter
is 1038, that of the white matter 1043. Therefore, the total weight of cortex
substance of both hemispheres would be 581 g, that of the white matter
approximately 464 g, and the total of both hemispheres 1045 g (MEYNERT states
1032 g). This corresponds closely to an average total brain weight of 1330 g,
whereby approximately 145 g is due to the cerebellum, and approximately 140 g
to the brainstem. Of course, the absolute volume of cortex gray matter
increases in the animal phylogeny upwards, despite the decrease in the
proportion to the white matter. According to CHR. JAKOB, the ratios of cortex
gray volume of the lower monkeys to the Orangutan and the human are as 1: 5:
24, since the increase of the whole cerebrum is so great. (The brain of a
full-grown Orangutan weighs approximately 500 g, with brainstem and
cerebellum.) JAKOB could not find a conspicuously regular difference between
right and left hemisphere cortical volumes. The left to right ratio was in one
case 290:250 cm3, but many times the cortex gray volume of the right hemisphere
was greater than that of the left.
Page 44
General remarks on the cortex and its neurons.
In
HENNEBERG's table the beautiful surface development of the Hottentott and Javan
brains are very notable, often surpassing the European brains - a warning
against deriving rushed conclusions from such data. WAGNER found 54,000 mm2 for
the surface of the full-grown orangutan brain, 21,000 mm2 of it free surface
and 33,000 mm2 hidden. CHR. JAKOB gives the ratio of the total cortical surface
of the monkey to the Orangutang to the human as 1:5:17.
which
we mentioned briefly in the last paragraph of chapter 1 (cf. p. 22). JAKOB
studied Gymnophiones [indeed Amphisbaenas, cf. Note
from Jakob's laboratory close to the beginning. MS], a type
which occupies a middle position between reptiles and amphibians. He showed
that the cells groups which first appear in the dorsal wall of the Gymnophion [Amphisbaenid: MS]
hemispheric pouch, and which later populate the cortex, are in continuous
contact with cells groups of the Corpus striatum. However, the top of the
Gymnophion [Amphisbaenid:
MS] cerebral pouch corresponds merely to Ammon's horn in
other animal species; otherwise, the cells layers correspond only to the
so-called inner main layer of the isocortex, i.e., layers V + VI. In Gymnophion
[Amphisbaenid:
MS] a new lamina develops from the rhinencephalon and
grows dorsally and finally medially over the lateral side of the hemisphere,
covering those cells which derive from the Corpus striatum (Fig. 16 a and b).
JAKOB correlates this phylogenetically later developing layer with the outer
main layer of the developed brain, while the lower layer corresponds to the
inner main layer. The outer "fundamental layer", as he calls it,
finds its phylogenetic origin in the rhinencephalon, and is continuous with
that outer cells layer. The inner "fundamental layer" derives
phylogenetically from the Corpus striatum, with whose lateral nuclear layer it
is connected. However, Ammon's horn is merely the direct continuation of this
inner fundamental layer. According to JAKOB, it completely lacks the outer
cells layer – as it does throughout the mammalian species, including humans.
Fig. 16a shows this relationship in Gymnophions [Amphisbaenas: MS].
In the embryonic development of the brain of the opossum JAKOB could also prove
this connection of the two fundamental layers with their original places, as
Fig. 16b shows. The two fundamental layers join together, and their joint
corresponds to the intermediate granular layer (our layer IV). This opinion has
much that speaks for it. The embryonic development, the ontogenesis of the
human brain no longer shows such a principal difference of the origins of the lower
versus the upper layers, since the anlage of the whole cortex is built by
migration of all neuroblasts from the underlying matrix. However, it is
possible that the ontogenetic development only imperfectly reflects the
phylogenetic stages, and that the migratory precursors of the inner and outer
layers already occur in the germinal anlage, skipping the intermediate stages.
If this brilliant opinion of JAKOB is really true, then it is correct for us to
identify the cells of part of the allocortex, that is Ammon's horn, with the
innermost two cells layers of the isocortex.
Structure
of the isocortex. Page 115
... We
will use these ratios for the wall thickness merely as comparison values, from
which one might in some cases find something noteworthy. For example: layer III
visibly loses thickness in the wall, yet, as seen above, gains relative width
from 33% at the cap to 37% on the wall, an increase of 4%. In certain areas of
the cortex, however, this difference is much greater; in the Gyrus rectus layer
III gains 10%, so that it even appears to narrow at the cap, rather than widen
as usual.
These
alterations of the layer thickness, which usually appear on the walls of each
gyrus, are not caused mechanically by the curvature of the gyral surface, but
they have quite special meanings due to the nature of each layer. The quite
colossal increase of layer VI at the cap, giving it a wealth of spindle cells,
and the nearly complete absence of layer VI and thus of spindle cells in the
valley, as well as the significant decrease of layer V from the cap to the
valley – together showing the reduction of the whole inner main layer in the
sulcal depths - certainly has a correlate in the different physiological
functions between cap, wall and valley (cf. this chapter, paragraph 5, p. 184).
Each individual gyrus becomes an individual organ, that consists of different
and differently structured parts, since every anatomical difference must of
necessity have certain physiological consequences. If one accepts this, then
the respective individual sizes, courses, connections with other gyri vía
so-called bridging gyri - in short the whole gyral architecture acquires a
quite different meaning than has been attributed to it until now. One cannot
consider it as a coincidence, as has recently and repeatedly happened due to an
overestimation of the micro-architectonic of the cortex, although the study of
gyral architectonics has unfortunately until now shown few positive successes.
Gyral structure leads us to suspect that it must have a special meaning, which
we do not yet know. If one considers the inner main layer as a primarily
efferent layer, like CHR. JAKOB (Chapter 2, cf. p. 22), and the outer as
receptive or associative, then this would throw some light on the meaning of
the structural differences between wall and cap. In future discussions on the
formation of the gyri one will have to take these factors into account, since
on this occasion they must play a crucial role. One sees the individuality of
these structural differences even where one does not think that it could really
be a wall formation because of further wall developments or possible secondary
gyral formations. For example, the Heschl gyri are only secondary transitional
gyri, between the parietal and temporal lobes, which lie on the dorsal surface
of the first temporal gyrus, the so-called Sylvian Surface. Throughout life
they carry the marks of a gyral wall in their general cellular structure and in
the narrowness of layers V and VI, which form a narrow stripe, even at their
caps. There are similar circumstances in the cuneo-parietal transition gyri at
the posterior wall of the upper parietal lobe in the Sulcus parietooccipitalis,
and elsewhere. Furthermore, it is probably no coincidence that in gyrencephalic
animals the so-called sensory cortex nearly always develops in gyral walls,
such as in the posterior wall of the Rolando Sulcus, in the dorsal wall of the
first temporal gyrus, in the wall of the Calcarina and in the interior wall of
the Gyrus cinguli. We know from above that the outer receptive main layer
prevails in the wall over the efferent layer, which decreases greatly here (cf.
also footnote p. 228).
Page 156
Details of the composition and meaning of the lamellar cortex structure.
Layer V
shows a quite particular behavior in the frontolimbic transition regions of the
median surface (Plates XVII, XXVI, XXXVIII, XXXIX), in the Gyrus rectus and in
the anterior part of the Gyrus cinguli (Plates XLV and XLVI), and especially in
the anterior Insula (Plate LIV). Here layer V or at least the upper part
attains such a density, through an increase of the cell numbers with good cell
sizes, that it forms a band through the cell picture of the cortex. Figs. 79-80
represent this approximately through the density of the hatching in a radial
direction. This formation is so conspicuous and clear in the anterior Insula
that we would like to call it the Insula belt, since it immediately
distinguishes the cortex of the anterior Insula, especially in thicker
sections. The significance of this "overdevelopment" as a cells band
is still quite uncertain. Maybe the corticofugal fibers to the thalamus
originate here, as such emanate especially amply from the median surface. This
conspicuous cells band in the upper part of layer V is located, as one sees, in
areas near the so-called rhinencephalon; in almost all such areas, other than
those quoted above, it is indicated but less intense, also in the edges of the
hippocampus (Plate CX). Besides this peculiarity, layer V shows some other peculiarities
at the cortex rim, to which we want to return later. In relation to the
"rhinencephalon" and its nearer and further surroundings, layer V
seems to play a not quite comprehensible role, in which apparently its
phylogenetically common origin with layer VI in CHR. JAKOB's (cf. p. 22) “inner
fundamental layer", the original and only lamina of the Archipallium (from
which Ammon's horn then develops), has a deeper meaning.
Page
l68 Details of the composition and meaning of the lamellar cortex structure.
This
leads us to the question about the relationships of layer V with the
allogenetic Cortex. ... Such pictures seem again to confirm BRODMANN's
assumption, who after detailed study, also at animals, thinks that merely layer
VI takes part in Ammon's horn. Maybe however this issue has less importance
than we today generally attribute to it. A cortex layer is not formed anyway at
this place in the embryonic brain (cf. p. 104 - 108), like as it occurs in the
Neocortex; therefore not either two fundamental layers, as CHR. JAKOB puts it,
or two steps, as LANDAU says and as they find in the anlage of the Neocortex.
But one single rung develops here; it is almost certainly that these
neuroblasts that form the Ammon's horn are closer by their nature to the cells of
the V than of those of VI, after which they form beautiful, big, very slender
pyramidal cells, as we otherwise are accustomed to see in V. Here, elucidation
must come from more detailed embryological and phylogenetic studies.
Structure of the isocortex Page
177
We have already mentioned in
Chapter 1, p. 22, the opinion of CHR. JAKOB that together the V + VI forms this
"inner fundamental layer"; LANDAU joins this opinion and calls them
"inner rung", and seems to assume that the whole "inner
rung" continues into the Ammon's horn, while as indicated BRODMANN assumes
this from VI alone.
Anyway,
layer VI is with the I the most constant in humans; it is excellently
developed, as we saw. Its upper part alone, VIa, amounts on average to 22% of
the cortical thickness! The whole VI is always the widest layer of the cortex,
with exception of quite certain places (for example the Koniocortex and
surroundings) and with the VIb usually amounts to virtually 40% of the actual
total cortical thickness in humans. We already saw earlier that some authors
mean that the width of V + VI grows as one goes further downward in the animal
row; against this, BRODMANN points out that some low mammals (small rodents and
insectivores) admittedly possess a notably wide layer of spindle cells (for
example rabbits), but on the other hand also superior mammals and the primates
exactly under the human level have a relatively very wide layer VI; furthermore
also some low clans have a very narrow layer VI (kangaroo). It therefore is not
generally right to say that the width of the most inner layer increases with
low animals! How it was already mentioned once, BRODMANN thinks also that one
could merely say that generally in the lower animals the inner main zone, IV +
V + VI, possesses in average a relatively larger width than the higher. In
order to be able to assess such circumstances, our tables should in the future
show also the corresponding relative ratios of the layers, i.e. their
proportions for homologous cortical locations in the animal row.
Structure
of Isocortex. Page 181
5.
Physiological meaning of the layers.
Although
our task is concerned with the bare morphology, yet at this place we must
touch, even if briefly, the question of the particular function of individual
layers. It probably is tightly connected with the question of the physiological
hierarchy of individual cells, about which we have already talked in the second
chapter section B, together with the description of the individual cells types;
and, furthermore, with the question of the fiber- and fibril architectonic,
which is outside our examinations. For that reason, by the sole means of
cytoarchitectonics we could not solve at all this question as we want; yet we
merely wish to remind of some possibilities that come from this study of
architectonic. Many times it was attempted to attribute determined functions to
the individual layers, an outlook which probably finds eloquent expression in
the sentence recently pronounced by VAN VALKENBURG, affirming that the cortex
consists of six joined peel-organs nicely imbricated in one another; also JAKOB
and VAN'T HOOG tend to a similar opinion. Against this opinion, it uses to be
remarked that the individual layers do not consist merely of a single cell type
each, but often from very different cells which probably have all their
particular significance. Additionally, one must consider that for example the
big- and giant-cellular layer V in the anterior central gyrus and the layer of
the smallest cells, V, in the parietobasal and occipital region present such a
different look that, if from cell composition a deduction regarding function is
at all admissible, one can hardly assume that the layers V can have one and the
same function in these two regions; so that therefore they do not represent the
same "organ". The same is applicable to the layer VI of these areas.
Nevertheless, even with full appreciation of these very justifiable objections
– which in any consideration of this question will never be allowed to forget –
one cannot still disregard that in the predominant part of the cortex the
molecular layer, the pyramidal layer, the granular layers, the ganglion cells
layer and the spindle cells layer constantly repeat themselves in a manner very
similar; and that, even if each of them is itself composed from several
individual cells types and cells layers, each layer is however composed to the
main part of only one cell type; this fact already finds expression in the
layer's name, too. One will therefore be also entitled, at least in a certain
sense, to search for a main function of the individual layers, whereby it can
become reputed, so that any other
function of the same layer may be usually held as additional, notwithstanding
that exceptionally some additional function could itself become the main one.
Very far, though, our present knowledge of this area do not yet lead us.
Structure
of the isocortex. Page 183
By reason
of phylogenetic studies, according to which layer III appears as the
phylogenetically most recent one, KAPPERS assumes that layer III serves the
higher interregional associations. Layer IV (granular layer) has receptive
functions, that serve infra-granular layers (V and VI), as origin of the
projection fibers as well as the intra-regional association.
CHR. JAKOB
has a similar opinion. We have already discussed in some detail his studies and
his conclusons (Chapter 1, p. 22 ff), and therefore refer again here on what
was said there, from which here we merely repeat: that the upper fundamental
layer (II + III) according to its opinion has above all a receptive (sensory)
function, the inner fundamental layer (V + VI) being mainly motor (efferent)
while the internal granular layer IV forms a system of short associations between
these two fundamental layers.
Through
isolation of the cortex of the hemisphere from the deeper centers, NISSL proved
with the old method of Gudden that, actually, only the inner layers V and VI
stands in connexion with the deep centers, thalamic nuclei, etc. With it, as
BRODMANN rightly says, a fundamental difference in function is proved between
the outer and the inner layers of the cortex' breadth.
FINES, on
reason of experimental sections of the corpus callosum, assumes that layer V
gives origin to the callosal fibers, while CAJAL, as mentioned, puts for it
layer IIIa.
As much
different as all these opinions firstly seem, yet they share many points of
contact. For example, that the V and VI are looked at as primarily effector
layers by all most recent authors. Therefore, it would also correspond that at
the sensory places of the brain, that is in the so-called koniocortex, the
layer V as well as the VI be quite weakly developed; and in fact specially the
first is sparsely populated, while the latter is specially narrow. On the
whole, behind the Rolandic sulcus layer V and furthermore layer VI do develop
less than in the forebrain where the motor functions are localized. In T2 and
T3, nevertheless, the V and VI do again nicely develop (from here after MONAKOW
the temporopontine tract should originate). However one cannot forget that also
the development of layer III has their optimum in the frontal brain!
To clarify
the role that the granular layer probably plays – which KAPPERS considers as
purely receptive and CAJAL (and CHR. JAKOB) as providing intracortical
connections between adjoining cortical areas and layers – we would like to add
the consideration that, as we saw in Chapter 4 (p. 150), layer IV can
admittedly consist of very different cells, all of which, probably, could
hardly have the same meaning. All the same, those small cells appear in
colossal amounts in the cortical areas that, as we still will see later,
represent centers of the sensory cortex, which therefore - in the koniocortex of
the centralis posterior, Heschl, calcarina, retrosplenialis and hippocampus –
present themselves simultaneously with an important increase (cf. p. 191) in
the exogenous fibers and the plexuses (in Va) from afferent fibers which (as
can be taken from CAJAL's impregnations) usually extend themselves under the
IV. This anatomical circumstance suggests that the small cells above all, and
specially those of the inner granular layer, must play an important role in the
receptive functions of the cortex; in particular, when they immediately receive
stimuli from the sensory fiber networks of Va, as already MEYNERT assumed it
too (1).
---
[footnote p 184] (1) That on occasions precisely in
the sensitive Koniocortex itself layer IV can be missing (for example, in hippocampal
granular HD), does not change anything at this opinion, since the other layers
are "granulated" at these places.
General anatomical discussion of the Areas.
Page 225
§2. Relationship of the areas' borders to the sulci
and gyri.
We
have just remarked that the areas' borders, in spite of their often being
chaotically laid by the course of sulci and gyri, many times crossing and
overlapping as already half a century ago BETZ did nicely know, in the frontal
lobe for example proceed horizontally, in such a way that the first and second
frontal sulci divide the forebrain into three big, horizontal frontal gyri,
pulling the Areas borders of FA, FB, FC, FD and FE a little backward and
downward (from front above to back below) and thus partition the cortex into
nearly sector-shaped Areas: compare Chapter 1, p. 23 CHR. JAKOB. On that
juncture the borders of the Areas against each other can just proceed crosswise
as well as alongside in the middle of a cap, or either the cap of a gyrus can be
covered by an Area while the gyral wall belongs to another Area. Furthermore,
as we want to discuss still later, the borders of these Areas show some
individual dislocations. This connection – seemingly altogether lacking for
some Areas – between tiling and cleavage of the cortex has recently led to a
strong underestimation of the meaning of the brain's gyral formations. First
and foremost, this remarkable paucity of connectionships between Areas and gyri
of the brain is not without exception; one can express this divergence by
saying that the sulci do not represent areal restrictions at all or do it only
in the rarest cases. In contrast, however, the localizations of a whole series
of well determined cortical formations are tied in to quite certain gyri or
sulci, their borders varying only in
small degree. This, for example, is applicable to the allocortex of the whole
so-called "rhinencephalon", which represents an individual and
peculiarly built formation, and to some extent also to the isocortex.
Page
226 Area division of the cortex.
The
anterior wall of the Rolando Sulcus, as well, and in ventral-dorsal direction
also increasingly the cap of the anterior central gyrus, is always the seat of
the Betz giant cells, the Area gigantopyramidalis, so that always the Rolando
fissure clearly separates two quite different architectural areas of the
Centralis anterior and Centralis posterior. Just as constantly, the
delimitation of the occipital and parietal cortex to the Sulcus
parietooccipitalis is at the median surface. … This points out that between
cortex construction and gyral construction, and probably not only with respect
to the primary fissures and constant sulci but, maybe, also regarding the
remaining sulci and gyri and their form, it must exist a not entirely opaque
connection, whose exposition is reserved to future research and in which CHR.
JAKOB's picture of the sector-shaped growth, that is the fan-shaped development
of the cortex on the one hand and the lengthwise folding (primordial-gyri
formation, Urgyrussbildung) on the
other hand, probably corresponds to its fundamental process (cf. p. 23).
Page
240 Area division of the cortex.
Besides
the above discussed granular Areas of the koniocortex, we have to look at the
more distant, specifically highly varied agranular Areas FA and FB situated
before the Rolando Sulcus. In these Areas, a transformation of most cells to
pyramidal cells takes place with nearly total loss of the granular layer, and
the whole area becomes characterized by a special size of the pyramidal cell;
even facing the whole remaining brain, the back part of the Area on the
anterior central gyrus is outstanding by reason of the well known development
of Betz giant cells. Already MEYNERT thought that this exceptional development
of the pyramidal cells was to be assumed as the expression for the motility,
and BETZ did the same since his giant cells' discovery, which cells he regarded
as specifically motor elements; the clinic and the experiment have generally
found this assumption right. However, the large pyramidal cells of the IIIc and
V layers, which one can regard as responsible for the motility, reach out over
the two agranular Areas: in point of fact, over the weakly granular Area FC
and, caudalward, the also granular part of FD, which we call FDm; as well as
over the whole third frontal gyrus until their orbitary part, which in the foot
already is by and large granular, namely FBCm, FDΓ, and FF. VOGT's new classic examinations, in which he
simultaneously took into account the architectural structure of this cortex and
the effect of its electric experimental stimulation, allowed him to ascertain
some alterations of the motor stimulation effects that go parallel with certain
alterations of the cortical cellular structure; a closer look of it we give
later, with the quite detailed discussion of the physiology of individual Areas
(Chapter 7, A, 3, §7, and Chapter 10, A, 5 §7). As it seems, one can say that
as immediate effect of stimulation the Area with giant pyramids arranges tonic special
movements, whereas FB primarily excites somewhat more severe tonic special
movements as well as so-called attitude movements (whole complexes). At any
rate, it is interesting to see in this example of the motility that a
qualitative alteration of the motor effect corresponds to an anatomical
alteration, and that the pyramid cortices FA and FB seem to be, specifically,
the motor efferent cortex. As well the leading Areas of large pyramidal cells,
FC, FDm, FDΓ, perhaps also FF, seem to have motor effects, besides others.
But motor effects from stimulation are also to be achieved from the posterior
central gyrus, Area PC; from the upper parietal lobe, Area PE; and elsewhere. A
point is to be taken for sure, anyway: pathology teaches us that human motility
depends in such a way on the forebrain that motility is not at all possible
without forebrain, while the monkey must, rather, be without cerebrum in order
to render motility not possible anymore. The progressive removal upward of the
sway of the neural axis' centers reached its maximum in the human and shows the
conquest of the whole motility by the cerebrum (JAKOB); in the colossal
development of the forebrain, this conquest expresses itself, both as regards
the regulation of the whole organism and in the particular control of the
motility of certain individual parts. One may compare the size of Areas FA, B,
C, D, Fig. 92; or fields 4, 6, 8, 9 of BRODMANN Fig. 6 in the human, with those
in the monkey, bear and rabbits on Fig. 100 and 101, 102 and 103, 104 and 105,
106 and 107 that are taken from BRODMANN's localizations.
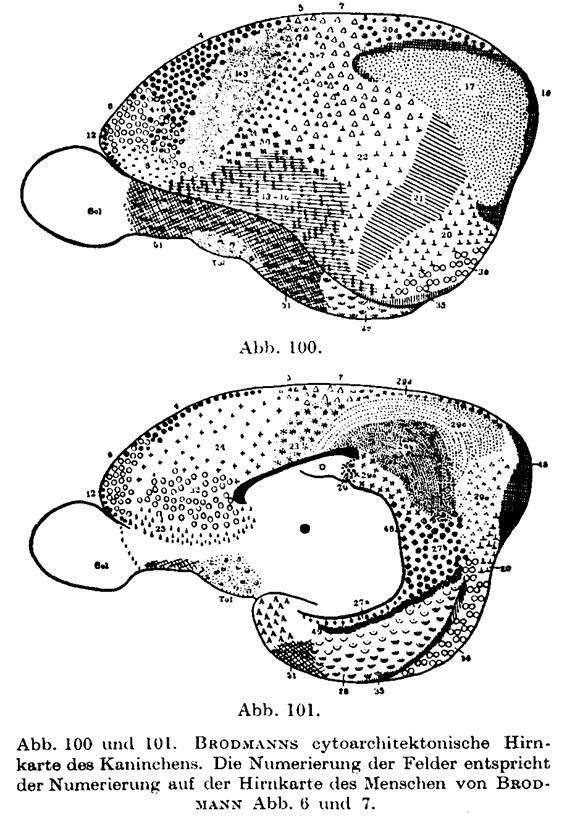
Fig.
100 and 101 [previous page]. BRODMANN's cytoarchitectonic brain map of the
rabbit. The numbering of the fields corresponds to the numbering on the brain
map of the human of BRODMANN Fig. 6 and 7.
Page
290 The forebrain.
Also
some pathological results speak in behalf of what we have already pointed out,
namely that the Betz giant cells primarily stand in the immediate context of
the motor function. So, in amyotrophic lateral sclerosis simultaneously with
the degeneration of the pyramidal tract one finds, also, a complete dwindling
of the Betz giant cells from the cortex (CAMPBELL, SCHRÖDER, ROUSSY and ROSSY).
Yet, we know from our own experience and from CAMPBELLs examinations that in
this pathology also the remaining pyramidal cells, especially those of layer
IIIc and the large pyramidal cells of layer V, are completely missing, and that
this atrophy of the Lamina ganglionaris can even encroach into the Area FB
frontal to it (SCHRÖDER, JAKOB, BUSCHER, among others). Later on, the atrophy
even encroaches on the cells of the IIIa and layer IIIb. Furthermore, CAMPBELL found a
change in the Betz giant cells of people to whom an extremity had been
amputated long time ago. He found these cells swollen, poor in branches, with
their Nissl bodies to some extent lost, and with the nucleus eccentrically laid
and no longer globular – an alteration that one must call reaction par distance; a particular decaying of
the cells that of course he did not find in amyotrophic lateral sclerosis.
Unfortunately CAMPBELL
omits, as he himself says, keeping an eye on the other
cells of the cortex in such amputation cases.
Area
praecentralis. Page 291
…
VOGT means with NISSL, CHR. JAKOB (cf. Chapter 1, p. 22), and FOERSTER among
others, that only the layers V and VI send the projection fibers out, whereas
as above said CAJAL as well as ourselves view the large pyramidal cells of the
IIIc and V layer as the places of origin of these fibers, at least as regards
the pyramidal tract. Still the first opinion is supported, in addition, by
BIELSCHOWSKY's, LENZ's and SPIELMEYER's examinations of the so-called
"paralyses with intact pyramidal tract". In these innate syndromes
the III layer of the cortex is nearly completely missing. These researchers
hence assume that layer III could not be the origin of the pyramidal tract, as
despite the big reduction in cell numbers that layer III offered in these cases,
the pyramidal tract seems anatomically intact; they rather assume that the
giant cells of layer V must mainly send the pyramidal tract fibers out, since
in these cases no anatomical changes have ever been pointed out in the
pyramidal tract itself. …
Page 336
The forebrain
1.
Area frontalis granularis FD (Plates XIX - XXX).
The
formation extends itself also from the Sulcus callosomarginalis at the median
brain wall over the edge to the median cortical surface and the convex surface
of the brain down into the Sylvian fissure and, frontalward, basal though
partially as far as into the orbitary surface of the forebrain. It therefore
forms on the whole another wide belt-shaped zone, that lies directly poleward
from the FC formation. The forebrain pole even remains free; it belongs with
another specific Area FE, nicely cut, that coats it with the approximate size
of a 5-Mark piece. Detached from this polar part the whole fore convex surface
of the forebrain (cf. Fig. 92) stands, therefore, occupied by FD; its rear
border coincides with the fore border of the FC, reaching farther to the front
to the first frontal gyrus; on the second frontal gyrus jumps back to the back
and on the third frontal gyrus falls between Pars triangularis and Pars
opercularis, thus approximately into the vertical branch of the Sylvian sulcus.
The fore border of the FD at the edge of the median cortical surface lies then
approximately 5 cm further poleward than the posterior part, and runs around
the pole forming a bow concave to the front that from here reaches as far as
the orbitary surface. In this way, the Area granularis frontalis from the back
to the front forms the fourth segment, which in frontal direction surrounds the
forebrain with a half-annular shape (FA, FB, FC, FD) (1).
---
[footnote p 336] (1) One compares this to CHR. JAKOB's sector pictures, that we
reproduce on Fig. 17 and 18.
Page
770 Lobus limbicus inferior.
…
So, as it becomes apparent from the cell forms too, one must take as likely
that as well as layer VI, also layer V has been tracked into the Subiculum's
pyramidal layer and is directly connected with it. Phylogenetically the opinion
seems even better vindicated that both layers of the inner main layer take part
in the formation of the Ammon gyrus; relating to this, JAKOB's genesis of the
fundamental layers can be seen in chapters 1, p. 20 - 24, as well as in the
following §6, p. 787. We have as well found the V layer connected with these allogenetic
formations also in the remaining allogenetic formations, for example at the
transition of the Area ultracingularis into Area obtecta (cf. p. 470).
Area
dentata. Page 787
According
to FLECHSIG, this area of the Uncus and the dorsal part of the hippocampal
gyrus belong to the early-myelinized primordial sensory centers, and he calls
it 4a and 4b. While he considers the Uncus to be part of the rhinencephalon, he
counts the Subiculum hippocampi and the posterior part of the gyrus fornicatus
(LE) to the taste sphere! - Physiologically the Uncus, because of its immediate
connection with the Tractus olfactorius, should not be assessed as cortex but
probably as ganglion.
We
previously mentioned that some researchers, BRODMANN among them, look at the Ammon's
horn as if it were a mere continuation of layer VI (p. 771); we quoted there
the reasons that speak for and against such an assumption. The phylogenetic
studies of ARIENS KAPPERS and CHR. JAKOB among others led to the realization
that the Ammon's horn with its single-cell layer is not any abortive cortex but
must be seen as a primitive cortex; in fact, as the very first cortex rudiment
(Fig. 15 and 16 a), which thrusts from the corpus striatum into the theretofore
membranaceous Pallium and remains on this primitive setting through the whole
animal kingdom and the whole lifespan through. KAPPERS therefore calls the
Ammon's horn 'Archipallium'. In contrast he names the remaining rhinencephalic
cortex (Uncus etc.) 'Paleocortex', so as to distinguish it from the
'Neocortex', which forms the whole remaining cerebral cortex. (We already
mentioned that the Neocortex approximately corresponds to our Isocortex
[homotypical plus heterotypical] and the Paleocortex and Archicortex to our
Allocortex.) The Archipallium (Archicortex) has purely a single rung (V and VI
= inner main layer = inner fundamental layer of CHR. JAKOB), the two other
cortex types have two cell rungs, that is the just named inner one and, onto
it, the outer fundamental layer. Now, LANDAU thinks that this outer rung is a
different one in the Paleocortex (Uncus) and in the Neocortex, and that in
longitudinal sections this outer main layer II and III of the Uncus clearly
distinguishes itself from the isocortex's II and III and also differs in the coloring.
Regarding the continuation of the structures of the Archipallium on the
Retrosplenium, LANDAU gets a similar opinion as we represented it on Fig. 129;
also he looks at the Taenia tecta and the Gyrus subcallosus as a continuation
of Ammon-cells layer (as we found LF1 of Area ultracingular posterioris, and
LF2 of Area obtecta as a continuation of HE1 and HE2). Furthermore, he says
that the cells of the Gyrus intralimbicus are the same as those of the Fascia
dentata, he being right inasmuch as the rear-Induseum's cells have the same
form as the granular cells of the Fascia dentata.

Prof. von Economo
_____
Copyright
© August 2005 del
autor / by the author. Esta es una investigación
de acceso público; su copia exacta y redistribución por cualquier medio están
permitidas bajo la condición de conservar esta noticia y la referencia completa
a su publicación incluyendo la URL original (ver arriba). / This is an Open
Access article: verbatim copying and redistribution of this article are
permitted in all media for any purpose, provided this notice is preserved along
with the article's full citation and original URL (above).

revista
Electroneurobiología
ISSN: 0328-0446
Croquis de este sitio -
Outline of this site
Índice - Table des matières - Inhaltsverzeichnis - Table
of Contents
Se recomienda mucho
leer o imprimir algunos de los trabajos mas extensos con el programa Acrobat
Reader,
que puede obtenerse gratuitamente pulsando aqui 
Haga doble "click" en el título de cualquier
artículo, para leerlo ahora - Double-click on any article to read it now:
*
SOCIOLOGÍA DE LAS NEUROCIENCIAS
¡Nuevo! "L’anthropologie ganglionnaire,
un psychovirus démasqué" (français)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF
(190 kB: recomendada) o .DOC
(76 kB).
*
ELECTRONEUROBIOLOGÍA
Efectos
relativísticos en biofísica cerebral:
An ESSENTIAL preprint: "Effects of relativistic motions
in the brain and their physiological bearing" (To be published in Helmut
Wautischer, ed., Ontology of Consciousness: A
Modern Synthesis) (English) (Tema: funcionamiento del cerebro y psiquismo)
Puede leer, imprimir o guardar en
su disco duro esta investigación en versión .PDF
(496 kB: recomendada) o .DOC (227 kB).
MBYKYHÁPE
GUARANÍME SUMARIO Y PÁRRAFOS INICIALES EN
CASTELLANO SUMÁRIO EM
PORTUGUÊS ABSTRAKTI
SUOMEKSI
*
Diversificación
de recursos electroneurobiológicos en la evolución del sistema nervioso:
Anticipo de impreso: "¡Alma e' reptil! Los
contenidos mentales de los reptiles y su procedencia filética" (Introducción a una
próxima edición de Biomedicina de reptiles por Juan Carlos Troiano y
colaboradores) (Castellano with a VERY LONG English abstract) (Tema:
origen del sistema nervioso)
Puede leer, imprimir o guardar en
su disco duro esta investigación en versión .PDF (733 kB: recomendada) o .DOC (406 kB).
SUMÁRIO EM
LÍNGUA PORTUGUESA
SUMARIO CASTELLANO
*
Cálculo
de potenciales dentro de las células
Calcule intensidades eléctricas y
magnéticas en cada compartimiento neuronal: "The nervous principle: active versus passive
electric processes in neurons"
(Explains how to calculate electric and magnetic field strengths inside
different neuronal compartments) (LONG FILE IN ENGLISH with
Bulgarian, Russian and Spanish abstracts/TOCs)
Podrá leer, imprimir o guardar en su disco duro
esta investigación en versión .PDF
(2 Mb): recomendada) o .DOC (1,5
Mb). También como .html comprimido (compressed .HTML folder: 0,5 Mb) .ZIP.
ENGLISH ABSTRACT AБСТРАКТ
НА БЪЛГАРСКИ
SUMARIO CASTELLANO
РЕЗЮМЕ НА
РОССИЙСКОМ
ЯЗЫКЕ
*
NOCIONES GENERALES
Conceptos:
 Noticia general -- ¿Qué es
electroneurobiología? -- La atmósfera intelectual (all in Spanish) -- Main Technical Ideas / Conceptos técnicos principales (English and Spanish) -- El descubrimiento de la Doppelrinde (German and Spanish)
Noticia general -- ¿Qué es
electroneurobiología? -- La atmósfera intelectual (all in Spanish) -- Main Technical Ideas / Conceptos técnicos principales (English and Spanish) -- El descubrimiento de la Doppelrinde (German and Spanish)
Historia de las
experimentaciones:
1883:
el primer mapeo con electricidad en todo el mundo ¡durante ocho meses! de un
cerebro humano consciente, realizado en San Nicolás, Provincia de Buenos Aires (Spanish)
Table of Contents (partial) of "Sensing: a new fundamental action
of nature" (English) -- Índices
Recepción de los aportes de Chr. Jakob en la
neurobiología germana
De "La citoarquitectonía de la
corteza cerebral" -- Constantin von ECONOMO y Georg KOSKINAS (1925) - Kommentar über die
Forschungsergebnissen von Prof. Christfried Jakob -
The comments on Professor Christfried Jakob's contributions made in 'The
Cytoarchitectonics of the Adult Human Cortex' by Professors Constantin, Baron
von Economo, and Georg N. Koskinas, rendered in English by Prof. H. Lee Seldon
(THIS PAGE); Los comentarios de Economo y Koskinas sobre las contribuciones de
Christofredo Jakob, traducidos por M. Szirko (Alemán, inglés, castellano:
páginas en construcción)
*
Panorama evolutivo:
Table of Contents of "Sensing: a new
fundamental action of nature" (English) -- Humour... -- Comment l’ hylozoïsme
scientifique contemporain aborde-t-il la sélection naturelle du parenchyme
neurocognitif? (français)
*
Vom Tierhirn zum Menschenhirn - Die
Organisation der grauen Substanz des menschlichen Zentralnervensystems (German and Spanish)
-- Christofredo Jakob: su vida y obra (Spanish)
*
FILOSOFÍA DE LA CIENCIA - CONCEPTO DE TIEMPO EN NEUROBIOFÍSICA
¡NEW! Why is Time
Frame-dependent in Relativity? Minkowski's spacetime as a Kantian 'condition of
possibility' for relativistic calculations (English) -- Abstract: Minkowski spacetime is a condition of
possibility for relativistic calculations. It keeps special relativity
kinematic (i.e. avoids introducing it it causal dynamics) but in exchange
forces relativistic observers to describe at once past, present, and future
states along the length of the observed moving entities. This has been
considered a proof that past and future components of real entities enjoy an
unremitting mode of existence, because, if the world described by relativity
were three-dimensional, the kinematic consequences of special relativity and
the experiments confirming them would be impossible. The latter is acknowledged
as exact but the assumed proof is viewed as incorrect because, such imposition
being a Kantian condition of possibility, it cannot reveal anything about what
Kant called noumenon, namely extramentality. ( Traducción castellana en preparación ).
You can
download a .PDF (1 MB:
recommended) or .DOC (571 kB) file for
printing this work.
*
“ANTAGONISMO ENTRE CIENCIAS DURAS Y HUMANIDADES BLANDAS”
H Markl: Dementia dichotoma: Der
Zweikulturenwahn - La ilusión de las dos culturas (Spanish and German) -- Vista del Sur: una nota a la American Association for
the Advancement of Science, Washington (Presidenta Jane Lubchenco) sobre
prioridades científicas (English)
*
MALFORMACIONES Y PAPEL DEL ÓRGANO CEREBRAL
Christofredo Jakob: “Los
Monstruos Anencéfalos” (Spanish)
Puede leer, imprimir o guardar en
su disco duro esta investigación en versión .PDF
(346 kB: recomendada) o .DOC (280 kB).
*
BIOÉTICA
¡Nuevo! Éthique de la Bio-Éthique (français)
Puede leer, imprimir o guardar en
su disco duro esta investigación en versión .PDF
(323 kB: recomendada) o .DOC (161 kB).
*
EL PRESUNTO DUALISMO CUERPO - ALMA
¡Nuevo! Soigner et Guérir ? Le rapport
soma-psyché et l’archéologie
du divorce culturel entre Médecine et Philosophie (français)
Puede leer, imprimir o guardar en su disco duro esta investigación
en versión .PDF (373 kB: recomendada) o .DOC (201 kB).
SUMARIO EN CASTELLANO
*
PSICOANÁLISIS Y FACILITACION PSICOSOMÁTICA DE LA ENF. DE
ALZHEIMER:
¡Nuevo! “CELOS:
Olvido, homosexualidad, desmemoria” [Temas: Celos y olvido. Los celos de los muertos. Celos y
homosexualidad. Los significados de ‘Untergang’. La ‘destrucción’ del complejo
de Edipo. Una hipótesis acerca de la desmemoria (destrucción de la memoria). La
enfermedad de Alzheimer.] (Spanish)
Puede leer, imprimir o guardar en su disco duro esta investigación
en versión .PDF (502 kB: recomendada) o .DOC (110 kB).
*
NUESTRA GENTE:
Reseña biográfica: Ramón Carrillo, el Gran Sanitarista
Argentino (Spanish)
*