Gobierno de la Ciudad de Buenos Aires
Hospital Neuropsiquiátrico "Dr. José Tiburcio Borda"
Laboratorio de Investigaciones
Electroneurobiológicas
y Revista
Еlectroneurobiología
ISSN:
0328-0446

Kommentar
über die
Forschungsergebnissen
von
Prof. Christfried Jakob
in

und
Dr. Georg N. Koskinas
em. Assistent der Psychiatrischen und
Neurologischen Universitätsklinik in Athen
Bearbeitet an der Psychiatrischen
Klinik
Hofrat J. Wagner v. Jauregg - Wien
(Wien und Berlin,
Verlag von Julius Springer 1925)
mit Englischen u.
Spanischen Übersetzungen
Englische
Übersetzung: H. Lee Seldon (Monash Univ., Australia)
wer auch den
ganzen deutschen Text mit 162 zum Teil farbigen Textabbildungen
und englischer
Übersetzung (in Bearbeitung) anbietet auf seinem Website http://neptune.netcomp.monash.edu.au/staff/lseldon/LeePublications.html
E-mail: Lee.Seldon[-at—]infotech.monash.edu.au
Spanische
Übersetzung: Mariela Szirko (Hosp. Borda, Buenos Aires: in Bearbeitung)
E-mail: MSzirko[-at—]Sion.com
DEUTSCHER TEXT: Electroneurobiología
2005; 13 (1), pp. 13 - 45; URL <http://electroneubio.secyt.gov.ar/index2.htm>
Copyright © Public.
This is a research work of public access, with redistribution granted on the
condition of conserving this notice in full and complete reference to its
publication including URLs. - Esta es una investigación de acceso público; su copia exacta y
redistribución por cualquier medio están permitidas bajo la condición de
conservar esta noticia y la referencia completa a su publicación incluyendo la
URL original (ver arriba).
*
Professoren von Economo und Koskinas machten ihre Kommentare (Literature, page 803), um sich auf CHR. JAKOBS Vom
Tierhirn zum Menschenhirn, vergleichend morphologische, histologische und
biologische Studien zur Entwicklung der Grosshirnhemisphären und ihrer Rinde,
I. Teil., München: F. J. Lehmann Vlg., 1911, und
Das Menschenhirn, Eine Studie über den Aufbau und die
Bedeutung seiner grauen Kerne und Rinde, München: F. J. Lehmann Vlg. 1911, zu beziehen.


Christfried Jakob (1866-1956) und
Constantin Freiherr von Economo (1876-1931)
--------
Allgemeiner Teil.
Allgemeine Grundlagen der Cytoarchitektonik der Großhirnrinde.
1.
Kapitel.
Vorbemerkungen.
A.
Einleitung.
Paarig, wie die Sinnesorgane, z.
B. die Augenblasen aus dem Zwischenhirn, so entwickeln
sich auch die Hemisphärenbläschen des Großhirns aus dem unpaarigen Endhirn, und man könnte die
daraus sich bildende Großhirnrinde als ein Sinnesorgan auffassen, dessen Blickfeld aber nicht auf die Aussenwelt, sondern
auf das innere Geschehen
im Zentralnervensystem selbst gerichtet ist. Die
Reize, welche dieses Organ treffen, kommen nicht
unmittelbar aus der Peripherie, sondern es sind bloß
Innenreize, die aus dem gesamten übrigen nervösen Organismus hierher gelangen,
er gemeinsam aufgefangen und zu einem Ganzen
verarbeitet werden. Die Großhirnrinde ist auch
imstande, diese Reizeindrücke anzusammeln, so daß jener überschüssige Teil der Reizenergie, der nicht im
einfachen Reflexbogen als direkter Effekt ich außen
wieder abgeleitet wird, sich in ihr staut. Dadurch befähigt, vergangene in gegenwärtige Energie in zukünftige umzuwandeln,
befreit sie den Organismus von dem brutalen primitiven Gesetz des Reflexaktes
und gibt ihm die individuelle Freiheit und die Persönlichkeit (CHR. JAKOB).

Professor Jakob seziert ein menschliches Gehirn auf dem sonnigen
Gelände seines Laboratoriums Südeingang (rechte Seite des Fotos). Zu der Zeit
(1906-7) entstand sein Interferenzmodell der widerhallende
(Resonanz) Makro- und Mikronetzwerken,
das in seiner Serie "Lokalisation der Seele und der Intelligenz"
erschien.
Historisches. S. 17
—
CHRISTFRIED JAKOBS Fundamentalschichten
20.
—
— Seine Urwindungen und
Sektorentheorie 23.
Drei Namen müssen noch erwähnt werden, die, wenn sie auch nicht direkt mit
der Cytoarchitektonik in Zusammenhang stehen, doch auf ihr weiteres Studium
stets von Einfluß bleiben werden, nämlich CAJAL, KAES und CHRISTFRIED JAKOB.
GOLGI hat uns mittels seiner
Silberimprägnationsmethode der Nervenzellen seit 1880 ein einzigartiges Mittel
in die Hand gegeben, die Form der Nervenzelle samt ihren Dendriten und ihrem
Achsenzylinder zu erkennen, und selbst zuerst damit uns grundlegende Kenntnisse
über die verschiedenen im Nervensystem vorkommenden Zellarten verschafft. Schon
bald darauf hat nun CAJAL sich systematisch an die Erforschung der menschlichen
und tierischen Großhirnrinde mittels der verschiedenen, auch von ihm selbst
wundervoll weiterentwickelten Silbermethoden gemacht, und unsere Kenntnisse, die
wir heute darüber besitzen, verdanken wir größtenteils diesem hochverdienten
spanischen Gelehrten. Im Laufe der Besprechung der einzelnen Zellformen im 2.
Kapitel (s. S. 44—68), ferner bei der Besprechung der einzelnen Areae und auch
sonst vielerorts kommen wir noch auf die einzelnen Befunde seiner ausgedehnten
Untersuchungen zurück. Die Kenntnis der gesamten Rindenarchitektonik kann uns
nur gemeinsam mit der Kenntnis des eben von CAJAL erforschten Baues der
einzelnen Zellelemente und der feineren Verbindungen derselben und des
Verlaufes der von ihnen abgehenden Bahnen zum richtigen Verständnis der im
Cortex ablaufenden Vorgänge bringen. Mit Bedauern müssen wir dabei die
Erfahrung registrieren, daß CAJALs verdienstvolle Arbeiten zum Teil ihrer Zeit
vorausgeeilt waren, da er dieselben anstellte, bevor die von MEYNERT und BETZ
postulierte areale Einteilung der Hirnrinde noch die nötigen gröberen
Grundlagen für die feineren Untersuchungen CAJALs geschaffen hatte. So ist es
heute oft schwer, die wichtigen Befunde, die in früherer Zeit mit der
Silbermethode gemacht wurden, genügend zu verwerten, weil die diesen feinen
Befunden entsprechen sollenden Stellen der Rinde nicht mehr ganz genau mit den
"Areae" homologisiert werden können. Daher hat sich CAJAL in unermüdlicher
Schaffenskraft neuerlich an die Silberimprägnation der einzelnen
"Areae" der Rinde herangemacht, und wir erwarten von diesen Studien
die wichtigsten Aufklärungen speziell auch für eine künftige
Fibrilloarchitektonik.
KAES hat 1907 Studien über die
normale (und pathologische) Hirnrinde mittels der Markscheidenmethode mit einem
beigegebenen Atlas veröffentlicht, deren wichtigste Ergebnisse wir hier kurz
zusammenfassen wollen: Die Rindendicke ist beim Neugeborenen in den ersten
Lebensmonaten bedeutender als später bei. Erwachsenen; vom dritten Lebensmonate
nimmt sie bis zum vollendeten ersten Lebensjahre progredient rapid ab, und von
hier geht die Abnahme langsam und progredient weiter bis zum Ende des 20.
Lebensjahres; um das 20. Jahr beginnt sie wieder zu steigen und erreicht im 5.
Lebensdezennium ihr Maximum, um dann wieder abzunehmen. Abb. 13, Kurve I, S. 21
gibt dieses Verhalten auszugsweise aus KAES' Originalbildern wieder. Die
Windungskuppe, die Windungswand und das Windungstal verhalten sich hierin
ziemlich gleichartig. Aber nicht alle Teile der Rinde machen diese Änderungen
ganz gleichmäßig mit. KAES teilt die Hirnrinde in eine äußere Lage, die bis zum
äußeren Baillargerschen Streifen reicht, also die äußeren drei Meynertschen
Schichten umfasst, die sog. äußere Hauptschicht und die darunterliegende innere
Hauptschicht. Nun zeigt KAES' Kurve II, Abb. 13, daß dieses wechselnde
Verhalten der Gesamtrindendicke speziell auf einer Dickenschwankung der äußeren
Hauptschicht beruht, die vorerst bis zum 20. Jahre abnimmt und dann bis zum 45.
bedeutend wieder wächst, während die innere Hauptschicht (Kurve III, Abb. 13)
eigentlich seit der Geburt progredient bis zum fünften Lebensdezennium sehr
langsam zunimmt. Sollte diese Feststellung sich als Regel bewahrheiten, so
kommt darin eine fundamentale Tatsache in der lebensphasisschen Entwicklung des
Gehirns zum Ausdruck, deren große Bedeutung sofort jedem einleuchtet. Für die
einzelnen Hirnteile behält nach KAES diese Kurve ihre Geltung in recht
verschiedenem Maße. Für das Stirnhirn B. gilt sie zur Ganze; für die Sehrinde
dagegen z.B. gilt sie nicht; die Entwicklungskurve zeigt hier keine großen
Schwankungen, sondern eine mehr kontinuierliche Entwicklung überhaupt. In
gewissen Hirnteilen verschiebt sich der Gipfel der Entwicklungskurve nach
anderen Lebensaltern; und so hat scheinbar jede Hirngegend ihre eigene Kurve,
betreffs welcher wir wohl auf die Originalarbeit KAES' verweisen müssen; auch
hier sind die regionalen Änderungen mehr durch das Verhalten der äußeren
Hauptschicht als der inneren Hauptschicht bedingt. KAES hat auch die Anzahl der
in der Hirnrinde in Millimeterbreite einstrahlenden Projektionsbündel in den
verschiedenen Lebensaltern bestimmt und eine Kurve dafür gegeben, die wir in
Abb. 14 auszugsweise wiedergeben und an der wir sehen, daß das Maximum um das
20. Lebensjahr erreicht wird; nun verschiebt sich sowohl die Anzahl der
Projektionsbündel als auch die Jahreszahl des Maximums regional auch wieder in
den einzelnen Teilen der Hirnrinde verschieden; am meisten weichen von dieser
Durchschnittskurve die Rinde der vorderen Zentralwindung und die Sehrinde ab.
KAES meint außerdem, die entwickeltere und faserreichere Rinde sei die
schmälere; beim Erwachsenen sei meist die Rinde der linken Seite die schmälere.
Wegen der besonderen Entwicklungsart und der verspäteten höchsten Entwicklung
der äußeren Hauptschicht (5. Dezennium!) glaubt KAES, daß dieselbe eine
besondere Rolle in der Entwicklung des individuellen und des höheren
Geisteslebens spiele. Gegen die Messungen KAES' hat man mit Recht eingewendet,
daß seine Zahlen — er gibt z. B. als Durchschnittsbreite der Rinde an den
Windungskuppen der Konvexität 4.9 mm anstatt recte höchstens 3.5 mm! — viel zu
hoch, also zum mindesten recht ungenau seien; und sicher wäre es sehr
wünschenswert, nachzukontrollieren, ob bei einer Richtigstellung derselben die
von KAES aufgestellten Regeln auch noch ihre Geltung behalten; denn es ist
zweifellos, daß wir für diesen Fall in diesen Regeln, besonders z. B. bezüglich
des Verhaltens der äußeren und inneren Hauptschicht, Sätze von fundamentaler
Wichtigkeit zu erblicken hätten. NISSL hat experimentell durch die Guddensche
Methode festgestellt, daß bloß die Zellen der inneren Hauptschicht in Beziehung
zu den tiefen Ganglien und Projektionsbahnen stehen; und auch diese Entdeckung
weist auf einen fundamentalen Unterschied zwischen äußerer und innerer
Hauptschicht hin. Wie sehr diese Schichten jede für sich dann regional ein
verschiedenes Verhalten aufweisen, werden wir im 4. Kapitel (s. S. 116—178)
sehen.

Professor von Economo, früh in seiner Beruf als Wissenschaftler
22 Vorbemerkungen.
CHRISTFRIED JAKOB hat nun in
seinen beiden noch unvollendeten Werken "Vom Tierhirn zum
Menschenhirn" und "Das Menschenhirn" ganz neue Forschungen veröffentlicht,
die, obschon eben falls wie die vorgenannten nicht unmittelbar mit der
Cytoarchitektonik in Zusammenhang, doch nicht ohne Einfluß auf den weiteren
Ausbau derselben bleiben können. Für die beiden Schichten der äußeren und der
inneren Hauptschicht, die er die beiden Fundamentalschichten der voll
entwickelten Rinde nennt, konnte er durch phylogenetische Studien und durch
seine Untersuchungen an Gymnophionen (Coecilis lumbricoides) — ein hierzu
besonders geeignetes Objekt, das betreffs seines Hirnbaues zwischen Amphibien
und Reptilien zu stehen scheint [Anmerkung von seinem Labor in 2005: einige Jahre später entdeckte
Jakob einen Fehler in der Taxonomie. Die mutmaßlichen Gymnophionen waren
tatsächlich Amphisbaeniden! Prof Jakob behandelte den Fehler mit Humor
während seines Lebens und berichtete darüber in einigen Büchern,
Artikeln und Briefen. Diese scheinen den Professoren von Economo und
Koskinas zur Zeit ihres eigenen Schreibens (1924) nicht bekannt gewesen
zu sein. Da die Systematik Jakobs Beobachtungen korrigiert wurde, hatte
der Fehler keine neurobiologischen Konsequenzen. MS] — einen verschiedenen Ursprung
für jede derselben nachweisen. Wir entnehmen seinem Buche die folgenden
Erklärungen und Abbildungen. Während das Großhirn bei Amphibien bloß aus dem
Riechhirn und dem Striatum besteht und die Decke des Hemisphärenbläschens, die
sich darüber spannt, noch rein ependymär ist (Abb. 15), zeigt sich bei der
Coecilia [Amphisbaena;
MS], wo diese Decke schon zu einem breiteren, Nervenzellen führenden
Gewebe entwickelt ist, daß die Nervenzellen dieser Formation (Archipallium),
die in der höheren Stufenleiter des Tierreiches dann zur Ammonsformation wird,
bloß der inneren Fundamentalschicht entsprechen und daß diese am seitlichen
Ansatz des Archipallium noch mit den Zellen des Striatum c. st. in
kontinuierlichem Zusammenhang bleiben (Abb. 16a, si). Von lateral her jedoch an
der Stelle (f.m.), wo sich in der Fissura marginalis das eigentliche Riechhirn
(Rh) absetzt, schiebt sich eine aus den Zellen dieses Riechhirns entspringende
Zellreihe (se) über die Zellen des Striatum und der inneren Fundamentalschicht
(si) nach außen darüber hinweg und bildet so den Ansatz zur äußeren
Fundamentalschicht (se), welche dann beide zusammen die gewöhnliche Rinde, das
Neopallium bilden. Bei embryologischen Studien über das Zentralnervensystem der
Beutelratten fand CHR. JAKOB Bilder, welche dafür zu sprechen scheinen, daß es
sich hier um ein allgemeingültiges Anlageprinzip handelt (Abb. 16b) (man
vergleiche hierzu die Abb. 66 Bild VI vom dreimonatigen menschlichen Foetus).
Er schließt daraus, daß die äußere (Hauptschicht) Fundamentalschicht
(II. + III. Schicht MEYNERTs) ihrem Ursprung aus dem Riechhirn nach prinzipiell
sensorischer, d. h. receptorischer Natur sei, während die innere Fundamentalschicht
(V. + VI.), die aus dem Striatum entsteht, motorischer, effektorischer Natur
sei; im späteren Leben vereinigen sich die beiden durch die IV. Schicht, deren
Körnerzellen ein System von kurzen Assoziationselementen zwischen den beiden
Fundamentalschichten bildet. Der Cortex der Urrinde, das Archipallium, bleibt
sich in seiner Entwicklung durch die ganze Tierreihe stets ziemlich ähnlich und
bildet die Ammonsformation; aus der lateralen Anlage mit den beiden
Fundamentalschichten dagegen wird das Neopallium (die eigentliche graue Rinde)
durch starkes Wachstum dorsal- und medialwärts und in die Breite; aus dem
Gebiete der Marginalfurche selbst wird die stets eigenartig gebaute Inselrinde
(mit dem Claustrum); außerdem nimmt das "Riechhirn" an der Basis
seine eigene weitere Entwicklung. Durch das starke Wachstum des Neopallium nun
von außen nach innen entwickelt sich die Rinde kolossal und legt sich in
Längsfalten, die Urwindungen, deren Innerste natürlich die
Ammonsrinde ist, dann der Gyrus limbicus und nach außen folgen, wie noch beim
Hundehirn als Urwindungen erkennbar, der Gyrus ectomarginalis, suprasylvicus,
ectosylvicus und der Gyrus insulae. Durch Überquellung des Rindenrandes an der
Fissura marginalis entsteht das Operculum. Neben dieser ventro-dorsalen
Entwicklung findet in frontocaudaler Richtung eine fächerförmig rotierende
Rindenentfaltung statt, deren Rotationspunkt im Inselgebiet liegt. Diese
Entwicklung bedingt (neben der obenerwähnten Segmentierung in Urgyri) einen
sektorenförmigen Bauplan in der Längsachse, der bei dem Cortexbau der
Lissencephalen noch deutlich zutage tritt. Abb. 17 zeigt dies deutlich (sie ist
CHR. JAKOB entnommen). Durch die Fortentwicklung und fächerförmige Entfaltung
in diesem rotierenden Sinne nach hinten entsteht dann der Occipitallappen und
durch Rückdrehung dieses hinteren Endes nach unten und wieder nach vorn der
Temporallappen, und so kommt das Sektorenbild der Primaten, das Abb. 18 nach
JAKOB wiedergibt, zustande. Jeder dieser
Sektoren hat seine physiologische Eigenart und eigene anatomische Verbindungen.
Ein Blick auf die Abb. 18 und auf unsere areale Hirnkarte (Abb. 19 und 20, die
wir hier zum Vergleiche in verkleinertem Maßstab wiedergeben) zeigt eine
gewisse frappierende Ähnlichkeit beider. Ebenso ein Vergleich der Abb. 17 mit
der arealen Hirnkarte der Lissencephalen Abb. 104 (S. 243). Ob nun diese
betreffs der Fundamentalschichten sowohl als auch der Sektorenentwicklung
neuartigen und grundlegenden Gedanken CHR. JAKOBs sich weiterhin als richtig
erweisen werden, wird die Zukunft zeigen; wir haben sie hier ausführlich
erwähnt, weil diese Auffassung der Hauptschichten zu unseren architektonischen
Studien jedenfalls in enger Beziehung steht und weil es ferner möglich ist, daß
die Ähnlichkeit der sektorenförmigen Entwicklung und der Grenzlinien der Areae,
die auf diesen Abbildungen sich zeigen, auf mehr als einer bloßen
Äusserlichkeit beruht.
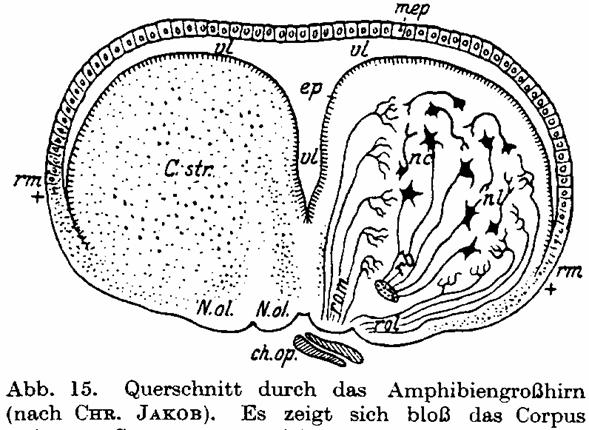
Abb. 15. Querschnitt durch das Amphibiengroßhirn (nach CHR. JAKOB). Es zeigt
sich bloß das Corpus striatum C.str. gut entwickelt, während die
Hemisphärendecke (Pallium) nur als dünnes ependymäres Häutchen den Ventrikel vl
dorsal abschließt mep.
Historisches. 23
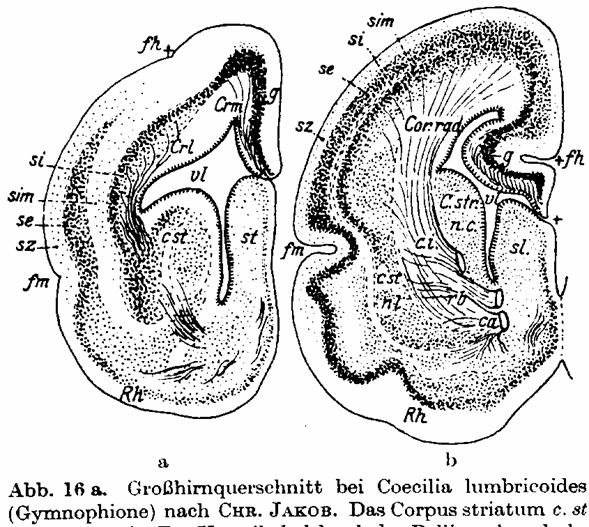
Abb. 16 a. Großhirnquerschnitt bei Coecilis lumbricoides (Gymnophione) nach
CHR. JAKOB.[Sie waren Amphisbaeniden, in der Tat. Siehe Anmerkung in den
"Geschichtlichen Notizen".] Das Corpus striatum c. st gut entwickelt. Der Ventrikel vl durch
das Pallium dorsal abgeschlossen, welches eine zwar dünne, aber doch schon
Nervenzellen führende Decke g (Archipallium) bildet. Diese Zellen entspringen
aus den lateralen Bandzellen des Corpus striatum und bilden das Stratum
internum si, die spätere innere Fundamentalschicht; Rh Riechhirn; fm Fissura
marginalis ist die Ansatzstelle des Riechhirns. Von hier aus zieht eine
Zellreihe se, die spätere äußere Fundamentalschicht von lateral und basal her,
dorsal über die si hinüber; die se stammt also ursprünglich aus dem Riechhirn
und verschmilzt später mit der si, die aus dem Striatum stammt, zum Cortex; sz
Stratum zonale, sim Stratum intermedium, fh Fissura hippocampi. — Abb. 16b
zeigt ähnliche Verhältnisse bei einem Großhirnquerschnitt eines Embryos der
Beutelratte. (CHR. JAKOB.)
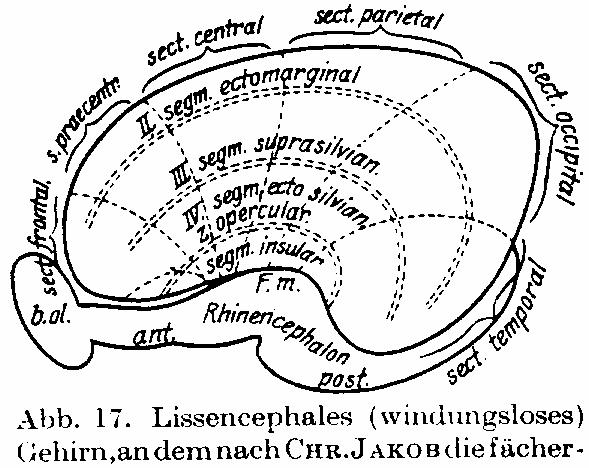
Abb. 17. Lissencephales (windungsloses) Gehirn, an dem nach CHR. JAKOB die
fächerförmige Entwicklung der Sektoren in frontocaudaler Entfaltung
eingezeichnet ist. Die Insel bildet gleichsam den Rotationspunkt dieser
Entfaltung. Auch die segmentale Gliederung ist eingezeichnet.
24 Vorbemerkungen.
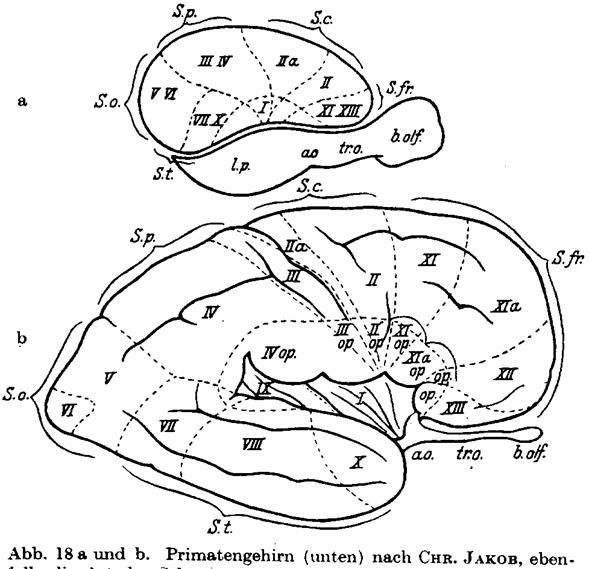
Abb. 18 a und b. Primatengehirn (unten) nach CHR. JAKOB, ebenfalls die Art
der Sektorenentwicklung bei einem hochentwickelten gyrencephalen Gehirn
darstellend. Durch die fächerförmige Drehung ist hier der Temporallappen
schließlich nach unten und vorn, und der Occipitallappen ganz nach hinten gerückt.
— Zum Vergleich ist darüber auch ein windungsloses Gehirn gezeichnet, um die
"Verschiebung" der Sektoren zur Anschauung zu bringen.
Während alle diese Studien,
denen sich noch die unserigen anschlössen, sich bemühten, die normale
Architektonik der Rinde aufzudecken, mit den verschiedenen hier aufgezählten
Zwecken und Aussichten, nicht zum wenigsten dem, die normale Grundlage zu
schaffen, die zur einwandfreien Erkennung pathologischer Veränderungen nötig
ist, hat die Untersuchung letzterer, ohne diese Resultate abzuwarten, man kann
sagen, so gut wie gleichzeitig ihren Gang genommen. Schon BETZ und HAMMARBERG haben, wie
gezeigt, Gehirne von. Idioten und KAES
daneben auch solche von Verbrechern untersucht. CAMPBELL und später SCHRÖDER
haben bei Pyramidenbahnläsionen und bei amyotrophischer Lateralsklerose
cytoarchitektonische Untersuchungen angestellt und KÖLPIN und LEWY haben bei
Huntingtonscher Chorea, SPIELMEYER und BIELSCHOWSKY bei Lähmungen ohne Läsion
der Pyramidenbahn, JOSEPH, A. JAKOB, BUSCAINO und KLARFELD, DOUTREBENTE und
MARCHAND bei Dementia praecox (Katatonie), ALZHEIMER, BRATZ, POLLACK, KOGERER
bei Epilepsie Veränderungen an Zellen und Schichten nachweisen können. C. und
O. VOGT haben in einer ausführlichen Abhandlung (Erkrankungen der Großhirnrinde)
die Grundlagen für eine künftige Pathoarchitektonik zu legen versucht, bei der
sie auch zahlreiche gute Bilder von normalen Rindenquerschnitten geben. Aus dem
Wiener neurologischen Institut von Prof. MARBURG sind mehrere ausführliche
Publikationen unter dessen Leitung und unter Mithilfe Dr. POLLAKs ausgeführt
worden, welche systematisch und eingehend die Pathoarchitektonik der Psychosen
behandeln, und zwar SAITO die progressive Paralyse, TAKASE das
manisch-depressive Irresein, NAITO die Schizophrenie und OSAKI die senilen
Psychosen. Schon früher hatte WADA in diesem Institut dieses Problem zum Objekt
seiner Studien gemacht. Der ersteren dieser Arbeiten ist als Grundlage zur
Beurteilung der pathologischen Veränderungen eine reiche Auswahl sehr guter photographischer
Bilder beinahe aller BRODMANNschen Felder mit einer bündigen und treffenden
Beschreibung derselben beigegeben. So sehen wir, mit welcher berechtigten und
gesunden Ungeduld schon allerorts die Pathologie eine erschöpfende Aufklärung
der normalen Verhältnisse der Hirnrinde erwartet.
Historisches. 25
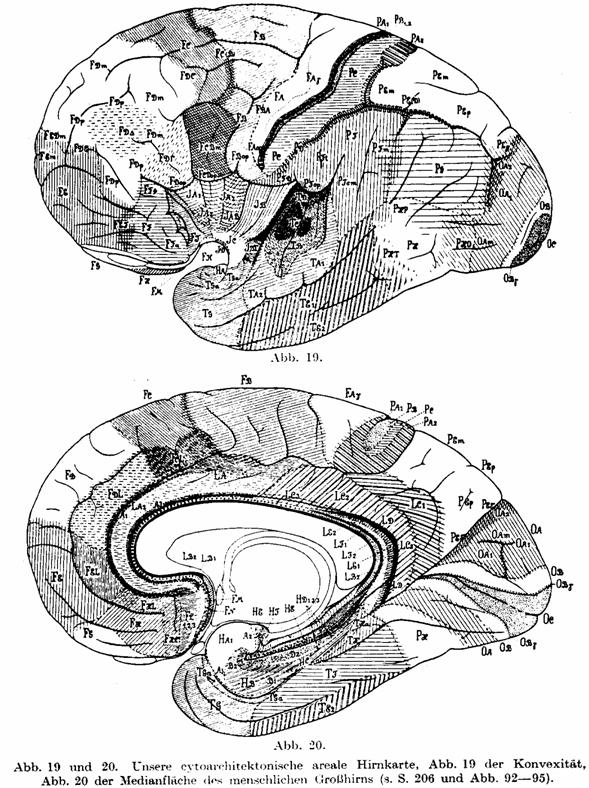
Abb. 19 und 20. Unsere cytoarchitektonische areale Hirnkarte, Abb. 19 der
Konvexität, Abb. 20 der Medianfläche des menschlichen Großhirns (s. S. 206 und
Abb. 92—95).
Rindenmaße. 41
3. Rindenvolumen.
Daß die graue Rinde in der Tierreihe aufwärts im Verhältnis zu
der Markmasse abnimmt, ist eine Tatsache, von der wir uns durch einen Blick z.
B. auf einen Hirnschnitt des Kaninchens überzeugen können (Abb. 31), bei dem
die graue Rinde kolossal breit ist und die Markmasse im Verhältnis zur
zellführenden Rinde nur einen ganz kleinen inneren Abschnitt bildet (vgl.
hierzu Abb. 25 der menschlichen Verhältnisse). Aber auch sogar beim Vergleich
vom Hirnquerschnitt eines niederen Affen, dann eines Orang-Utan und eines
Menschen sieht man diese progressive Zunahme an Markmasse und relative Abnahme
und Verdünnung der Rinde bis zum Menschen. Bei niederen Affen überwiegt nach
CHR. JAKOB die graue Substanz über die weiße am Querschnitt noch im Verhältnis
von 5:1; beim Orang nur mehr im Verhältnis 3:1, beim Menschen ca. 2:1.
42 Allgemeines
über den Cortex und seine Nervenzellen.
JAEGER hat das Volumen der grauen Rinde der Hemisphären
bestimmt und zum Volumen des Markes in Proportion gebracht. Er bestimmte das
Volumen an Hirnscheiben bestimmter Dicke mittels der von Anton angegebenen
planimetrischen Messungsart und errechnete das Volumen der grauen Rindenmasse
beider Hemisphären zusammen auf 540—580 cm3, das der weißen Masse auf 400—490
cm3 (ohne Stamm); im Durchschnitt wäre also das Volumverhältnis von Rindengrau
zu Markweiß 560:445 oder ca. 1.2: 1. Die Dichte der grauen Substanz ist nach
DANILEWSKI 1038, die der weißen 1043. Also ist daraus zu errechnen das
Gesamtgewicht der grauen Rindensubstanz beider Hemisphären mit 581 g, das der
weißen Markmasse mit ca. 464 g, das Totalgewicht beider Hemisphären mit 1045 g
(MEYNERT gibt 1032 g an), was bei einem mittleren Gesamthirngewicht von 1330 g
dem richtigen Verhältnis ziemlich genau entspricht, da doch ca. 145 g davon
aufs Kleinhirn, ca. 140 g auf den Hirnstamm entfallen. Natürlich ist das
absolute Rindengrauvolumen, trotz der Abnahme in der Proportion zum Markweiß,
in der Tierreihe aufwärts ein immer größeres; so verhält sich nach CHR. JAKOB
das Rindengrauvolumen des niederen Äffen zum Orang und zum Menschen wie 1: 5:
24, da die Zunahme des ganzen Großhirns eine so bedeutende ist. (Das Gehirn des
ausgewachsenen Orangs wiegt ca. 500 g, mit Hirnstamm und Kleinhirn.) Einen
auffallenden regelmäßigen Unterschied zwischen rechter und linker Hemisphäre
konnte JAKOB betreffs des Rindenvolumens anscheinend nicht finden; zwar
verhielt sich die linke zur rechten einmal wie 290:250 cm3, aber wiederholt war
das Rindengrauvolumen der rechten Hemisphäre auch wieder größer als das der
linken.
44 Allgemeines
über den Cortex und seine Nervenzellen.
Sehr auffallend nun in der
HENNEBERGschen Tabelle ist die schöne Oberflächenentfaltung der Hottentotten-
und Javanergehirne, welche vielfach das Europäerhirn übertreffen — eine
Warnung, aus solchen Befunden übereilte Schlüsse zu ziehen.
WAGNER fand für die Oberfläche des ausgewachsenen
Orang-Utan-Gehirns nur die Zahl 54000 mm2, davon 21000 mm2 freie Oberfläche und
33000 mm2 verborgene; und CHR. JAKOB gibt als Verhältnis der Gesamtoberfläche
des niederen Affen zum Orang, zum Menschen die Gleichung an 1:5:17.
106 Aufbau
von Entwicklungsgrundlagen der lamellären Cortexgliederung.
Man sieht ferner aus den Bildern
der Abb. 66, daß schon recht früh auch der embryonale Übergang vom Neocortex
mit seiner deutlich gebildeten Pyramidenschicht zum Allocortex mit seinen
Anlagen aus lockeren Zellverbänden zwar ein ziemlich rascher ist, daß aber die
Grenze (a und b) keine absolut fixe ist und daß es auch Übergangsbildungen an
diesen Grenzen gibt. So ist es auch beim Erwachsenen zwar möglich, zwischen dem
Bau des Isocortex und dem des Allocortex genau zu unterscheiden, jedoch sind
auch hier die Grenzgebiete zwischen beiden einerseits vom isogenetischen Cortex
recht verschieden, andererseits wieder weisen sie vielfach deutlich genug noch
immer die sechs Schichten auf, um ihre Einteilung kurzweg zum Allocortex etwas
zu erschweren, so daß man oft im Zweifel ist, zu welcher Bildung man so eine
Grenzformation zählen soll. Der Allocortex striatus, den wir eben früher
erwähnten, bildet gewöhnlich eine solche Grenzformation und in der embryonalen
Entwicklungsart desselben finden wir auch die Begründung für diese
Zwitterstellung. Wir werden bei der späteren Besprechung der Areae sehen, daß
diese Abgrenzung im Gebiete des Uncus, des übrigen Hippocampus, der
retrosplenialen Teile des Gyrus cinguli und im Gebiete der Area parolfactoria
und auch an anderen Stellen oft Schwierigkeiten bietet. Mit Rücksicht auf das
eben Gesagte ist es wohl auch ein undankbares Unternehmen, wenn man versucht,
die Zellagen, welche später einmal den vollentwickelten Allocortex bilden, mit
den gewöhnlichen sechs Schichten des Isocortex oder einem Teile derselben zu
identifizieren, da doch die Anlage schon eine recht verschiedene ist und der
spätere scheinbare Zusammenhang vielleicht doch nur ein rein äußerlicher ist.
Allerdings haben hier in letzter Zeit bedeutungsvolle Untersuchungen von CHR.
JAKOB ganz neue Gesichtspunkte eröffnet, auf die wir schon im letzten Absatz
des 1. Kapitels (s. S. 22) flüchtig eingegangen sind. JAKOB hat seine
Untersuchungen an Gymnophionen gemacht [tatsächlich
Amphisbaeniden, cf. Siehe Anmerkung vom
Jakobs Labor in den
"Geschichtlichen Notizen". MS], einer Gattung, welche zwischen
Reptilien und Amphibien eine Art Mittelstellung einnehmen. Hier konnte er nun
nachweisen, daß die in der dorsalen Wand des Hemispharenbläschens der
Gymnophionen [Amphisbaena: MS] zuerst
auftretenden Zellverbände, welche die Rinde bevölkern, noch in kontinuierlichem
Zusammenhang mit den Zellverbänden des Corpus striatum sind. Die Decke des
Hemisphärenbläschens der Gymnophionen [Amphisbaeniden:
MS] entspricht aber bloß der Ammonsformation der übrigen Tierreihe;
außerdem entspricht diese Zellage aber bloß der sog. inneren Hauptschicht der
Rindenschichten des Isocortex, d.h. bloß der V. + VI. Schicht; denn man sieht
eben bei diesen Gymnophionen [Amphisbaena: MS]
an der lateralen Seite der Hemisphäre aus dem Riechhirn heraus eine neue
Zellage sich entwickeln und sich über die eben erwähnte Zellage der
Hemisphärendecke, welche aus dem Corpus striatum stammt, dorsalwärts und medialwärts
hinüberschieben (Abb. 16 a und b). Diese sich phylogenetisch
später entwickelnde, aus dem Riechhirn stammende äußere Zellage homologisiert JAKOB mit der äußeren Hauptschicht des
entwickelten Gehirns, während die untere Lage der inneren Hauptschicht entspricht. Die
äußere "Fundamentalschicht", wie er sie nennt, findet also ihren phylogenetischen Ursprung im Riechapparat, in dessen
äußere Zellschicht sie sich
auch fortsetzt, die innere "Fundamentalschicht" ist phylogenetisch
aus dem Corpus striatum hervorgegangen, mit dessen lateraler Kernschicht sie
streckenweise noch zusammenhängt. Die Ammonsformation ist aber die direkte
Fortsetzung bloß dieser inneren Fundamentalschicht, Sie entbehrt nach JAKOB
vollständig des äußeren Zellstratums, ein Verhalten, das sie in der ganzen
Säugetierreihe auch weiterhin bis zum Menschen charakterisiert. Abb. 16a gibt
dieses Verhältnis bei Gymnophionen [Amphisbaeniden: MS]
wieder. In der embryonalen Entwicklung des Gehirns der Beutelratte hat JAKOB,
wie Abb. 16b zeigt, diesen Zusammenhang der beiden Fundamentalschichten mit
ihren Ursprungsstellen scheinbar auch noch nachweisen können. Die beiden
Fundamentalschichten verbinden sich miteinander und ihre Verbindungsstelle
entspricht der Zwischenkörnerschicht (unserer IV Schicht), Diese Auffassung hat
etwas sehr Bestechendes für sich. Die embryonale Entwicklung, die Ontogenese am
Menschenhirn zeigt nun allerdings nicht mehr eine solche prinzipielle
Verschiedenheit der Entstehung der unteren Schichten von den oberen, da die
Anlage der ganzen Rindenschicht durch eine Auswanderung aller Neuroblasten aus
der darunterliegenden Matrix entsteht. Jedoch wäre es sehr möglich, daß die
ontogenetische Entwicklung hier die phylogenetischen Stadien nur unvollkommen
wiedergibt und daß schon in der Keimanlage die für die spätere Entwicklung
nötigen Verschiebungen mit Überspringung von Zwischenstadien stattfinden.
Trifft diese geniale Ansicht JAKOBs wirklich
zu, dann ist es allerdings richtig, daß wir wenigstens für einen Teil des
Allocortex, und zwar für die Ammonsformation die Zellen derselben bloß mit den
Zellen der Innersten zwei Zellagen des Isocortex identifizieren können.
Bau des Isocortex. 115
Was wir nun schon früher bezüglich der absoluten Werte für die Windungswand
gesagt haben, daß ihnen nämlich die Fehlermöglichkeit anhaftet, daß nicht immer
ganz gleichwertige Stellen gemessen wurden, da es recht schwer ist, gerade die
Mitte der Windungswand anzugeben, gilt in noch erhöhtem Maße vielleicht für
diese relativen Zahlen. Wir werden also diesen Verhältniszahlen für die
Wanddicken weniger Bedeutung beimessen und werden dieselben bloß als Vergleichswerte
benutzen, aus denen man fallweise einiges Beachtenswerte manchmal doch
ermitteln kann. Als Beispiel hierfür möge folgendes angeführt sein: es nimmt
die III. Schicht in der Wand, wie gesagt, an Dicke immer sichtlich ab; ihr
relativer Wert jedoch nimmt, wie aus obigen Verhältniszahlen hervorgeht, da die
übrigen Rindenschichten sich viel rascher und in viel bedeutenderem Maße in der
Wand verschmälern, von 33% an der Kuppe bis auf 37%, also um volle 4% zu. In
gewissen Teilen des Großhirns jedoch ist diese Differenz eine viel bedeutendere
und nimmt die III. Schicht z. B. im Gyrus rectus sogar um 10% zu, d. h. daß sie
sich an der Kuppe sogar scheinbar verschmälert, statt wie gewöhnlich im
Verhältnis zur Wand sich zu verbreitern.
Diese Änderungen der Schichtendicke,
die gewöhnlich an jeder Windung in ihren Wänden wiederkehren, sind nicht etwa
mechanisch durch die Krümmung der Windungsoberfläche bedingt, sondern sie haben
ihre ganz besondere, dem Wesen jeder Schicht anhaftende Bedeutung. Die ganz
kolossale Zunahme der VI. Schicht am Culmen, die einen ganz besonderen Reichtum
der Windungskuppe an Spindelzellen bedingt, und das beinahe vollkommene Fehlen
der VI. Schicht, also der Spindelzellen im Tal, sowie die bedeutende Abnahme
der V. Schicht im Verhältnis vom Culmen zum Tal, womit also auch die ganze
innere Hauptschicht in den tiefen Windungspartien bedeutend reduziert wird, hat
sicher ihr Korrelat in einer Differenz der physiologischen Funktion zwischen
Kuppe, Wand und Tal (vgl. dieses Kapitel, Abs. 5, S. 184). Dadurch wird jede
einzelne Windung eigentlich zu einem eigenen Organ, das aus verschiedenen und
verschieden gebauten Teilen besteht, da doch jede anatomische Verschiedenheit
notwendig auch gewisse physiologische Folgen haben muß. Betrachtet man aber in
diesem Sinne jede Windung als ein Organ, dann gewinnt die jeweilige
individuelle Größe derselben, ihr Verlauf, ihre Zusammenhänge mit anderen
Windungen durch sog. Brückenwindungen, kurz die ganze Windungsarchitektonik
eine ganz andere Bedeutung, als man ihr bis jetzt zugeschrieben hat. Man kann
diese nicht mehr als eine Zufälligkeit ansehen, wie es in einer Überschätzung
der Mikroarchitektonik der Hirnrinde in den letzten Jahren wiederholt geschehen
ist, obschon allerdings bisher das Studium der Windungsarchitektonik leider nur
wenig positive Erfolge aufzuweisen hatte. Der Aufbau der Windung jedoch läßt
uns vermuten, daß derselbe eine ganz besondere Bedeutung haben muß, die wir
heute allerdings noch nicht genau kennen. Fasst man die innere Hauptschicht im
Sinne CHR. JAKOBs (2. Kap., s. S. 22) als hauptsächlich effektorische Schicht
auf, die äußere als receptorisch oder assoziativ, so würde schon diese
Auffassung einiges Licht auf die Bedeutung dieser Bauunterschiede zwischen Wand
und Kuppe werfen, und man wird wohl auch in Hinkunft bei den Diskussionen über
die Entstehung der Windungen dieses Moment nicht mehr unberücksichtigt lassen
dürfen, da es entschieden hierbei eine ausschlaggebende Rolle spielen muß. Man
sieht die Eigentümlichkeit dieses Bauunterschiedes der einzelnen Teile des
Windungsquerschnittes auch dort, wo man wegen einer weiteren Entwicklung der
Wand und eventueller sekundärer Windungsbildungen auf ihr gar nicht mehr denkt,
daß es sich wirklich um eine Wandbildung handeln könnte. So sind z. B. die
Heschlschen Windungen nichts anderes als sekundäre Übergangswindungen vom
Parietallappen zum Temporallappen, welche sich auf die dorsale Wandfläche, die
sog. Sylvische Fläche, der ersten Temporalwindung erstrecken, und sie tragen in
ihrem allgemeinen Zellbau und in der Schmalheit ihrer V. und VI. Schicht, welch
letztere gar bloß ein schmales Band bildet, zeitlebens das Wandgepräge, sogar
auch an der Windungskuppe der Heschlschen Windungen. Ähnliche Verhältnisse
findet man auch an den cuneo-parietalen Übergangswindungen an der hinteren Wand
des oberen Parietallappens im Sulcus parietooccipitalis und auch anderwärts.
Ferner ist es dementsprechend wohl auch kein Zufall, daß bei Gyrencephalen der
sog. sensorische Cortex hauptsächlich und beinahe immer in Windungswänden sich
entwickelt, so in der Hinterwand der Rolandoschen Furche, in der Dorsalwand der
ersten Temporalwindung, in der Wand der Calcarina, in der Innenwand der Gyr.
cinguli; und wir wissen aus dem oben Gesagten, daß die äußere receptorische
Hauptschicht in der Wand überwiegt über die effektorische, welche hier stark
abnimmt (s. auch S. 228, Fußnote).
156 Einzelheiten
über die Zusammensetzung und die Bedeutung des lamellären Rindenbaues.
Ein ganz besonderes Verhalten
weist ferner die V. Schicht in den frontolimbischen Übergangsteilen der
Medianfläche (Tafel XVII, XXVI, XXXVIII, XXXIX), auch im Gyrus rectus und im
vorderen Teil des Gyrus cinguli (Tafel XLV und XLVI) und ganz speziell in der
vorderen Insel (Tafel LIV) auf, wo durch Zunahme der Zellzahl bei guter
Zellgröße die V. Schicht oder wenigstens ihre obere Lage eine solche
Zelldichtigkeit erlangt, daß sie bandartig das Zellbild der Rinde durchzieht.
Abb. 79, 80 stellt dies durch die Dichtigkeit der Schraffierung in radiärer
Richtung ungefähr dar. Besonders in der vorderen Insel ist diese Bildung so
auffallend und deutlich, daß wir sie hier als Inselgürtel bezeichnen möchten,
da daran allein schon die Rinde der vorderen Insel an etwas dickeren Schnitten
ohne weiteres immer erkennbar ist. Was diese "Überentwicklung" als
Zellband zu bedeuten hat, ist noch ganz unbestimmt. Vielleicht nehmen hier die
corticofugalen Fasern zum Thalamus ihren Ursprung, welche ja von der
Medianfläche besonders reichlich entspringen sollen. Dieses auffallende Zellband
in der oberen Lage der V. Schicht findet sich, wie man also sieht, in Gebieten
in der Nähe des sog. Riechhirns; etwas weniger scharf als in den angeführten
Teilen findet man es zum mindesten angedeutet auch in beinahe allen übrigen
gegen das Riechhirn abfallenden Rindenpartien, auch in den Randpartien des
Hippocampus (Tafel CX). Ausser dieser Eigentümlichkeit zeigt die V. noch einige
andere Zelleigentümlichkeiten am Rindensaum, auf die wir alle später nochmals
zurückkommen wollen. Die V. Schicht scheint ja überhaupt in bezug auf das
"Riechhirn" und seine nähere und weitere Umgebung irgendeine
besondere, uns noch nicht ganz verständliche Rolle zu spielen, bei der offenbar
ihre phylogenetisch gemeinsame Anlage mit der VI. Schicht zu der von CHR. JAKOB
(s. S. 22) "innere Fundamentalschicht" genannten ursprünglichen und
einzigen Zellage des Archipalliums (aus der sich dann die Ammonsbildung
entwickelt) irgendeine tiefere Bedeutung hat.
168 Einzelheiten
über die Zusammensetzung und die Bedeutung des lamellären Rindenbaues.
Dies führt uns zur Frage über
die Beziehungen der V. Schicht zum allogenetischen Cortex. Daß auf die Teile
des allogenetischen Cortex, die man Cortex striatus nennt, alle Rindenschichten
mit Ausnahme der IV. übergehen können, haben wir bei Besprechung der I., II.
und III. schon angedeutet, es geht auch die V. Schicht auf sie über, z. B. in
der Uncusformation (Tafel C und CI, CV usw.). Auch auf den Cortex
rudimentarius, als den man wohl das auf den Balkenrücken sich umschlagende
Rindenstückchen (des Gyrus intralimbicus) bezeichnen muß, setzt sich, wie auch
aus Tafel XL, XLVII, L und LII mit großer Wahrscheinlichkeit zu entnehmen ist,
die V. Schicht fort und nimmt an seinem Zellaufbau teil; großenteils scheint
mir allerdings die Va nicht mehr auf den Balkenrücken zu gelangen, sondern eher
bloß die schlanken Zellen aus der Tiefe von Vb. Nun ist dieser lamelläre Cortex
auf dem Balken (Gyrus intralimbicus) zum Teil nur eine Fortsetzung der
Ammonsformation. Aber es ist, wie mir scheint, heute noch unentschieden, ob die
V. Schicht sich auf die Rinde des Cortex rudimentarius der Ammonshornformation
tatsächlich fortsetzt oder ob bloß die VI. Schicht hinüberzieht. Bilder, wie
sie Tafel CII3 und CVII—CXII geben, ließen es als wahrscheinlich erscheinen,
daß die großen Pyramidenzellen des rudimentären Cortex der Ammonsformation HE
mit den überschlanken Pyramidenzellen der V. Schicht der Nachbarschaft HD
direkt in Zusammenhang stehen, während nur die dünnen Lagen tiefster und meist
schon horizontal gestellter spindelförmiger Zellen unterhalb dieser schönen
Pyramidenzellen des Ammonshorns die Fortsetzung der VI. Schicht zu sein
scheinen. Andererseits sieht man wieder Bilder wie Tafel CV (Uncusgegend), auf
welchen die V. Schicht in der oberen Wand der mittleren Windung 2 des Uncus am
Kuppenwinkel anscheinend endet; unter ihr scheint dann bloß die VI. Schicht
ihre Fortsetzung zu finden und in weitem Bogen in die Kuppe 1, der Endigung der
Ammonsformation im Gyrus digitatus unci, einzuziehen und schließlich mit den
Zellen des Cortex primitivus der Kuppe 1', die bloß ans einer Molekularschicht
und aus Zellen des N. amygdalae besteht, zu verschmelzen. Solche Bilder
scheinen wieder BRODMANNs annahme zu bestätigen, der nach eingehendem Studium
auch an Tieren meint, daß bloß die VI. Schicht an der Ammonsformation
teilnehme. Vielleicht hat aber diese Frage weniger Bedeutung, als wir ihr heute
im allgemeinen beimessen. Eine Rindenschicht wie im Neocortex wird an dieser
Stelle im embryonalen Hirn jedenfalls nicht gebildet (s. S. 104—108), also
jedenfalls auch nicht zwei Fundamentalschichten, wie CHR. JAKOB, oder zwei
Staffeln, wie LANDAU sich ausdrückt und wie sie sich in der Anlage des
Neocortex finden. Sondern es entwickelt sich hier bloß eine einzige Staffel;
daß diese Neuroblasten nun, die die Ammonsformation bilden, ihrem Wesen nach
den Zellen der V. näherstehen als denen der VI., nachdem sie schöne, große,
überschlanke Pyramidenzellen bilden, wie wir sie sonst in der V. zu sehen
gewohnt sind, ist höchstwahrscheinlich. Hier müssen eingehendere embryologische
und phylogenetische Studien Licht bringen. (Über die Auffassung, daß die
subiculäre und die Ammonsformation eine Nebeneinanderschaltung von
zusammengehörigen Schichten darstellen, statt einer Übereinanderschaltung, wie
wir sie sonst im Cortex finden, also gleichsam eine Verzerrung der
Hirnschichten in horizontaler Richtung, darüber wollen wir im speziellen Teil
im Kapitel 13 über die hippocampischen Bildungen sprechen.)
Bau des Isocortex. 177
Auch dem allogenetischen Cortex
zu ändert die VI. etwas ihre Zellzusammensetzung, und zwar erfolgt das meist
derart, daß ihre oberste Lage Dreiecksform annimmt, die untere Lage aber
unbestimmt meist horizontal oder eigentümlich schief gestellte, manchmal
gestreckte, manchmal gekrümmte, kahnartige oder gewundene Formen zeigt, die die
ursprüngliche Spindelform nur noch erraten lassen; diese letzteren Zellen sind
meist spärlicher, verlieren sich allmählich im Mark und bilden keine deutliche
Schicht, wie in der isogenetischen Rinde; derart verändert hängt die VI.
Schicht mit den Zellen des allogenetischen Cortex zusammen. Wenn es über die
anderen Schichten zweifelhaft ist, ob sie sich auf den Allocortex fortsetzen
oder nicht — wir verweisen diesbezüglich auf das S. 139 und 168 Gesagte —, so ist
die VI. Schicht wenigstens jedenfalls ohne weiteres in sichtbarem Zusammenhange
damit. Im Allocortex allerdings haben die Zellen nicht mehr
Spindelzellencharakter, sondern einen sehr verschiedenen, und die Schicht
verbreitert sich manchmal ganz bedeutend, so z. B. im Ammonshorn, wo sie aus
ganz großen Pyramidenzellen besteht, die eine ganz dichte zellreiche und
zellgroße Schicht bilden, an deren untersten Saum bloß einige horizontale
kleine Spindelzellen noch zu sehen sind. Mancherorts kann diese dichte Pyramidenschicht,
mit der I. überkleidet, allein ganze Gyri bilden, z. B. am caudalen Teile des
fimbriären Ende des Uncus, wo sie die sog. Gyri digitati bildet. Aber
diesbezüglich verweisen wir auf den speziellen Teil und die Beschreibungen der
Areae des Hippocampus 13. Kap, und Tafel CV, CVI.
Wir haben schon im 1. Kap., S.
22, die Ansicht CHR. JAKOBs erwähnt, daß die V + VI zusammen die ..innere
Fundamentalschicht" bilden, und LANDAU schließt sich dieser Ansicht an und
nennt sie "innere Staffel", und scheint anzunehmen, daß die ganze
"innere Staffel" sich in die Ammonsformation fortsetzt, während
BRODMANN dies, wie gesagt, von der VI allein annimmt.
Jedenfalls ist die VI. Schicht
mit der I. die konstanteste beim Menschen; sie ist, wie wir gesehen haben, vorzüglich
entwickelt. Ihr oberer Teil allein (VIa) beträgt durchschnittlich 22% der
Rindendicke! Die ganze VI. aber ist immer die breiteste Schicht der Rinde, mit
Ausnahme ganz bestimmter Stellen (z. B. Koniocortex und Umgebung) und beträgt
mit der VIb gewöhnlich nahezu gegen 40% der eigentlichen Gesamtrindendicke beim
Menschen. Wir haben nun früher schon gesehen, daß einige Autoren meinen, die
Breite der (V +) VI wachse, je weiter man in der Tierreihe nach abwärts gehe;
BRODMANN sagt dagegen, daß zwar manche niedrigstehende Mammalier (kleine Nager
und Insektenfresser) eine auffallend breite Spindelzellenschicht besitzen (z.
B. Kaninchen), andererseits hätten aber auch hochstehende Säuger und unter den
Primaten gerade der Mensch eine relativ sehr breite VI Schicht; außerdem haben
auch manche tiefstehende Sippen eine sehr schmale VI Schicht (Känguruh). Es sei
also nicht generell richtig, zu sagen, daß die Breite der innersten Schicht bei
niederen Tieren zunehme! Wie schon früher einmal gesagt, meint auch BRODMANN,
man könne im allgemeinen bloß sagen, daß bei niederen Tieren die innere
Hauptzone (IV + V + VI) eine relativ- größere durchschnittliche Breite besitze
als bei höheren. Um solche Verhältnisse überhaupt bewerten zu können, wird man
in Zukunft unseren Tabellen entsprechend die relativen Verhältniszahlen der
Schichten, d.h. ihre Proportionalgleichung für homologe Rindenstellen auch in
der Tierreihe bestimmen müssen.
Bau des Isocortex. 181
5. Physiologische Bedeutung der
Schichten.
Obwohl bloß die Morphologie
unsere Aufgabe ist, müssen wir doch an dieser Stelle, wenn auch nur ganz kurz,
die Frage einer bestimmten Funktion der einzelnen Schichten streifen. Sie ist
wohl eng verbunden mit der Frage nach der physiologischen Dignität der
einzelnen Zellen, auf die wir schon im 2. Kap., B, jeweilig bei Beschreibung
der einzelnen Zellarten zu sprechen gekommen sind, ferner mit der Frage der
Faser- und Fibrillenarchitektonik, die ausserhalb unserer Untersuchungen liegt.
Wir wollen und können also noch gar nicht an Hand der Cytoarchitektonik allein
diese Frage lösen, sondern wollen bloß versuchen, auf einige Möglichkeiten, die
sich aus dem Studium der Architektonik ergeben, hinzuweisen. Es ist schon
vielfach versucht worden, den einzelnen Schichten bestimmte Funktionen
zuzuschreiben, und diese Ansicht findet wohl ihren beredten Ausdruck in dem
Satze, die Rinde bestehe aus sechs schalenartig ineinander gefügten Organen,
welche neuerlich von VAN VALKENBURG ausgesprochen worden ist; auch JAKOB und
VAN'T HOOG neigen zu einer ähnlichen Auffassung. Gegen diese Ansicht wird mit
Recht geltend gemacht, daß die einzelnen Schichten nicht bloß aus je einer
einzigen Zellart bestehen, sondern oft aus sehr verschiedenen Zellen, die
wahrscheinlich alle ihre eigene Bedeutung haben; ferner muß man sich auch
überlegen, daß z. B. die groß- und riesenzellige V Schicht in der vorderen
Zentralwindung und die kleinstzellige V Schicht in der parietobasalen und
occipitalen Region ein so verschiedenes Aussehen haben, daß, wenn ein Schluß
aus der Zellzusammensetzung auf die Funktion überhaupt statthaft ist, man kaum
annehmen kann, daß die V. Schicht in diesen beiden Regionen überhaupt ein und
dieselbe Funktion haben kann, also auch nicht das gleiche "Organ"
darstellt. Dasselbe gilt für die VI. Schicht dieser Gegenden. Bei aller
Würdigung dieser sehr berechtigten Einwände, auf die man bei einer Überlegung
dieser Frage nie wird vergessen dürfen, darf man aber doch wieder nicht ausser
acht lassen, daß im überwiegenden Teil der Hirnrinde die Molekularschicht, die
Pyramidenschicht, die Körnerschichten, die ganglionäre Schicht und die
Spindelzellenschicht doch in sehr ähnlicher Weise sich immer wiederholen und
daß, wenn sie auch jede einzelne aus mehreren Zellgattungen und Zellagen sich
zusammensetzen, sie doch zum Hauptteil aus je einer Zellart bestehen, die ja
auch im Namen der Schicht schon ihren Ausdruck findet; man wird also auch in
gewissem Sinne berechtigt sein, wenigstens nach einer Hauptfunktion der
einzelnen Schichten zu fahnden, wobei es als gegeben angesehen werden kann, daß
neben dieser Funktion noch die eine oder andere Funktion derselben für
gewöhnlich zukommen kann, ja sogar in Ausnahmefällen diese Nebenfunktion selbst
zur Hauptfunktion werden könnte. Sehr weit führen uns heute allerdings unsere
Kenntnisse auf diesem Gebiete noch nicht.
Bau des Isocortex. 183
KAPPERS nimmt auf Grund
phylogenetischer Studien, denen zufolge die III. Schicht sich als die
phylogenetisch jüngste ergibt, an, daß die III. Schicht den höheren
interregionalen Assoziationen diene. Die IV. Schicht (Körnerschicht) habe
rezeptorische Funktionen, Die infragranulären Schichten (V und VI) dienen als
Ursprung der Projektionsfasern und der intraregionalen Assoziation.
CHR. JAKOB hat eine ähnliche
Auffassung. Wir haben seine Studien und seine Schlüsse (1. Kapitel, S. 22)
etwas eingehender schon besprochen und verweisen daher hier nochmals auf das
dort Gesagte, von dem wir hier bloß wiederholen: daß nach seiner Ansicht die
obere Fundamentalschicht (II + III) vor allem eine rezeptorische (sensorische)
Funktion hat, die innere Fundamentalschicht (V + VI) prinzipiell motorisch
(effektorisch) ist, die Zwischenkörnerschicht IV aber ein System von kurzen
Assoziationen zwischen diesen beiden Fundamentalschichten bildet.
NISSL hat in Anlehnung an die
alte Guddensche Methode durch Isolierung des Cortex der Hemisphäre von den
tieferen Zentren nachweisen können, daß tatsächlich nur die inneren Schichten V
und VI im Zusammenhang mit den tiefen Zentren (Thalamuskernen usw.) stehen. Damit
ist, wie BRODMANN richtig sagt, ein fundamentaler Funktionsunterschied zwischen
den äußeren und den inneren Lagen des Rindenquerschnittes erwiesen.
FINES nimmt auf Grund
experimenteller Balkendurchschneidungen an, daß die V. Schicht den Balkenfasern
ihren Ursprung gebe, während CAJAL, wie oben erwähnt, die IIIa-Schicht dafür
anspricht.
So verschieden auch alle diese
Ansichten für den ersten Augenblick zu sein scheinen, so berühren sie sich doch
andererseits wieder in vielen Punkten, so z. B. darin, daß die V. und VI. als
hauptsächlich effektorische Schichten von allen neueren Autoren angesehen
werden. Dem entspräche es auch, daß an den sensorischen Stellen des Gehirns,
das ist also im sog. Koniocortex, die V. Schicht sowohl als die VI. ganz
schwach entwickelt sind, und zwar die erstere besonders zellarm, die letztere
besonders schmal. Im ganzen hinter der Rolandoschen Furche gelegenen Gehirn ist
ferner die V. und VI. weniger entwickelt als im vorderen Gehirn, wo die
motorischen Funktionen lokalisiert sind. In T2 und T3 sind aber die V. und VI.
wieder gut entwickelt (hier soll nach MONAKOW die temporopontine Bahn
entspringen). Allerdings darf man nicht vergessen, daß auch die Entwicklung der
III. Schicht im Frontalhirn ihr Optimum hat!
Zur Erklärung der Rolle, die
die Körnerschicht wahrscheinlich spielt und die KAPPERS als eine rein
rezeptorische, CAJAL (und CHR. JAKOB) als eine intracortical nächstgelegene
Rindenteile und Schichten miteinander verbindende ansieht, möchten wir die
Überlegung hinzufügen, daß zwar die IV. Schicht, wie wir im 4. Kap. (S. 150)
gesehen haben, aus sehr verschiedenen Zellen bestehen kann, die wohl kaum alle
dieselbe Bedeutung haben können, daß aber so kleine Zellen in kolossalem Maße
in jenen Rindenpartien auftreten, welche, wie wir später noch sehen werden,
sensorische Rindenzentren darstellen — also im Koniocortex der Centralis
posterior, Heschl, Calcarina, Retrosplenialis und Hippocampus — gleichzeitig
mit einer bedeutenden Zunahme (s. S. 191) des unter der IV. für gewöhnlich sich
ausbreitenden exogenen Fasere nd geflochtes (in Va) von zuführenden Fasern, wie
aus CAJALs Imprägnationen entnommen werden kann. Dieser anatomische Umstand
legt wohl den Gedanken nahe, daß die kleinen Zellen überhaupt und speziell die
der inneren Körnerschicht doch bei den rezeptorischen Funktionen der Rinde, und
zwar beim unmittelbaren Auffangen des Reizes aus den sensorischen
Fasergeflechten der Va eine wichtige Rolle spielen müssen, wie es auch schon
MEYNERT annahm 1).
[footnote p 184 1. Daß dabei im sensiblen
Koniocortex selbst manchmal die IV. Schicht gerade fehlen kann (z. B. in
hippocampischen granulösen HD), ändert nichts an dieser Auffassung, da an
diesen Stellen die anderen Schichten "verkörnelt" sind.]
Allgemein-Anatomisches über die Areae. 225
§ 2. Beziehung der arealen Grenzen zu
den Furchen und Windungen.
Wir haben gerade vorher gesagt,
daß die arealen Grenzen oft, unbekümmert um den Verlauf von Furchen und
Windungen, dieselben vielfach überkreuzend, dahinziehen, was schon BETZ vor einem
halben Jahrhundert bekannt war; während z. B. im Frontallappen die erste und
zweite Frontalfurche horizontal verlaufen und das Stirnhirn in die drei großen,
horizontal ziehenden Stirnwindungen teilen, ziehen die arealen Grenzen von FA,
FB, FC, FD und FE etwas geneigt von vorne oben nach hinten unten und teilen die
Rinde beinahe sektorenförmig ein (vgl. 1. Kap., S. 23 CHR. JAKOB). Die Grenzen
der Areae gegeneinander können dabei ebenso gut quer als längs mitten auf einer
Kuppe verlaufen, oder es kann die Kuppe einer Windung mit einer Area bedeckt
sein und die Wand der Windung schon zu einer anderen Area gehören. Die Grenzen
dieser Areae zeigen außerdem, wie wir später noch besprechen wollen, einige
individuelle Verschiebungen. Dieser anscheinend für einige Gegenden vollkommen
fehlende Zusammenhang zwischen Felderung und Furchung der Rinde hat in letzter
Zeit zu einer starken Unterschätzung der Bedeutung der Windungsbildungen des
Gehirns geführt. Vor allem gilt dieser anscheinende Mangel jeder Beziehung zwischen
Areae und Windungen des Gehirns nicht durchweg und man kann diese Diskrepanz
bloß so richtig zum Ausdruck bringen, indem man sagt, daß die Furchen keine
arealen Begrenzungen darstellen oder nur in den seltensten Fällen. Andererseits
sind jedoch eine ganze Reihe von ganz bestimmten Rindenbildungen an ganz
bestimmte Windungen oder Furchen in ihrer Lokalisation gebunden, und es können
ihre Grenzen nur in geringem Grade variieren. Das gilt z. B. vom Allocortex des
ganzen sog. "Riechhirns", welches eine eigene und eigenartig gebaute
Bildung darstellt, zum Teil aber auch für den Isocortex. So fand schon WERNICKE
an der medianen Hirnfläche die Wände der Calcarina stets mit der typischen
Formation, die wir heute Koniocortex der Area striata (OC) nennen, ausgekleidet,
welche, caudalwärts keilförmig zunehmend, auch auf die dorsale, cuneale und
ventrale, linguale Lippe der Calcarina heraustritt und am Occipitalpol etwas
auf die Konvexität bis in die Nähe des sog. Sulcus lunatus (auf den Gyrus
descendens Ecker) übergreift; das Ausmaß, in welchem diese Bildung auf die
Lippe der Calcarina und die Konvexität übergreift, ist individuell verschieden,
die Tatsache jedoch, daß die Area Oe die Calcarinawände einnimmt, ist konstant.
Auch die übrigen "Koniocortex" sind, wie schon einmal erwähnt (s. 4.
Kap., S. 116), an die Wände bestimmter Sulci gebunden, so z. B. der Koniocortex
der Retrosplenialgegend an die Aussenwand des caudalen Teiles des Sulcus
callosus, d.h. an die Innenwand des Gyrus limbicus (Pars posterior), und er
tritt nur in der Isthmusgegend auf das Culmen des Gyrus; der Koniocortex des
Allocortex, d. h. des Praesubiculums des Gyrus hippocampi, ist an die Kante
zwischen Kuppe und Wand des Gyrus hippocampi in den Sulcus hippocampi und
dessen ganze frontocaudale Ausdehnung gebunden und greift bald weiter auf das
Culmen des Gyrus, bald tiefer in die Wand des Sulcus über; der Koniocortex der
hinteren Zentralwindung ist stets in der Vorderwand derselben, d. h. in der
hinteren Wand des Sulcus Rolando, und erstreckt sich hier von der opercularen
Region des Sulcus bis dorsal zu seinem parazentralen Ende und tritt erst an der
Mantelkante auf die Oberfläche des Parazentralläppchen heraus; der Koniocortex
der ersten Temporalwindung findet sich immer auf der dorsalen Wand derselben,
und zwar auf deren Nebenwindungen, den sog. Heschlschen Windungen.
226 Areale
Einteilung des Cortex.
Ebenso ist die Vorderwand der
Rolandoschen Furche und in ventral-dorsaler Richtung auch zunehmend die Kuppe
der vorderen Zentralwindung stets der Sitz der Betzschen Riesenzellen, der Area
gigantopyramidalis, so daß die Rolandosche Furche immer deutlich zwei ganz
differente architektonische Gebiete der Centralis anterior und Centralis
posterior trennt. Ebenso konstant ist an der Medianfläche die Abgrenzung von
der Occipital- und Parietalrinde durch den Sulcus parietooccipitalis.
Weniger genau lokalisiert, doch
ebenfalls konstant sind auf der dritten Frontalwindung der Reihe nach die Area
Broca (FCBm auf der Pars opercularis), die Area triangularis (FDΔ auf der
Pars triangularis) und die Area praetriangularis FFΦ vor derselben und die
FF auf der Pars orbitalis der dritten Frontalwindung und die Area FI auf der
vorderen orbitobasalen Wand der Insel und dem Gyrus transversus insulae. Doch
läßt sich allerdings mit solch annähernder Genauigkeit der Sitz und die
Ausdehnung nicht aller Areae bestimmen. Jedenfalls bestehen gewisse Beziehungen
zwischen einzelnen eigenartigen Cortexbildungen und den primären großen
Fissuren; dies gilt besonders für jene, welche schon früh zur Anlage des
Allocortex in Beziehung stehen, die Fissura hippocampi, Fissura marginalis,
Fissura rhinica (Abb. 56—58), und ferner zeigen einige konstante Furchen
konstante Beziehungen zu der Felderung, hierzu gehören die Calcarina, Parietooccipitalis,
Rolando. Aus diesen Grundlagen läßt sich dann ungefähr die Lage auch der
übrigen Areae bestimmen; allerdings scheinen hier beim ersten Anblick die
inkonstanteren, sekundären Furchen mit der Felderung nichts zu tun zu haben.
Daneben bestehen noch zwischen
der Ausbreitung gewisser Areae und der Größe und Form der Windungen ebenfalls
gewisse Beziehungen, die wir heute noch nicht genauer fassen können. So ist es
z. B. für das Frontalhirn (s. S. 316) nachweisbar, daß die dickrindigen Areae
FA, FB und FC auf großen, runden, kuppelartig gewölbten Windungen sich
ausbreiten, während die Area FD weiter frontal immer erst dort beginnt, wo die
Windungen auf eine mittlere Schmalheit herabsinken, das kleinzellige FE jedoch
und FDp, der kleinzellige Teil der FD-Rinde, breiten sich auf schmalen,
kleinen, stark gewundenen Gyri aus, und je weiter caudalwärts in einem Gehirn
die Verschmälerung der Windungen im Frontallappen beginnt, eine desto größere
Ausdehnung caudalwärts nimmt auch die Area FE vom Frontalpol nach hinten. Ein
ähnliches Verhältnis zwischen Windungsbau und Rindenbau läßt sich auch an
anderen Hirnstellen finden, z. B. zeigt am hinteren Ende des Parietalhirns die
Schmalheit der Windungen den Beginn der Occipitalformation an. Zwischen der Art
der Ausbreitung der Area LA am Gyrus limbicus und der Form des Sulcus
callosomarginalis besteht ebenfalls eine noch ungeklärte Beziehung; s. S. 444.
Dies alles weist darauf hin, daß zwischen Rindenbau und Windungsbau wohl nicht
nur bezüglich der primären Fissuren und konstanten Furchen, sondern vielleicht
auch betreffs der übrigen Furchen und Windungen und ihrer Form ein noch nicht
näher eruierbarer Zusammenhang bestehen muß, dessen Erschließung künftiger
Forschung vorbehalten bleibt und bei dem CHR. JAKOBs ansicht des
sektorenförmigen Wachstums, d. h. der fächerförmigen Entfaltung der Rinde
einerseits und der Längsfältelung (Urwindungsbildung) andererseits
wahrscheinlich einem grundlegenden Vorgang dabei entspricht (s. S. 23). Daneben
ist es sehr gut möglich, da doch die Rinde das primär Wachsende ist und da die
Differenzierung der Felder Gebiete nicht nur verschiedener Dicke und
verschiedenen Aufbaues, sondern wohl auch verschiedener Wachstumsenergie
hervorbringt, daß dadurch, wie WETZEL sagt, Spannungserscheinungen entstehen,
welche die Bildung von Windungen und Furchen mitbedingen; und es spricht
wirklich nicht gegen einen solchen Zusammenhang, daß die Grenzen der Areae
nicht mit den Furchen zusammenfallen, denn auch die rein mechanischen Beugungen
in einer Platte, die aus verschieden biegsamen Teilplatten zusammengesetzt
wäre, würden nach diesem Autor nicht genau an den Übergangsstellen derselben,
sondern daneben erfolgen — ganz abgesehen davon aber sind hier jedenfalls nicht
mechanische Momente, sondern Wachstumsvorgänge das Primäre. Alle die
vorgebrachten Argumente zeigen jedenfalls das Bestehen von teilweise schon
bekannten, andererseits wenigstens in Umrissen sich zeigenden neuen
Zusammenhängen zwischen Windungsbildung und Architektonik, bei der auch noch ganz
andere Kräfte im Spiele sind (s. S. 115).
240 Areale
Einteilung des Cortex.
Ausser den oben besprochenen
granulösen Areae des Koniocortex haben wir als fernere, spezifisch
hochdifferenzierte Areae die vor der Rolandoschen Furche gelegenen agranulären
Areae FA und FB anzusehen, in welchen bei beinahe völligem Verlust der
Körnerschicht eine Umwandlung der meisten Zellen zu Pyramidenzellen stattfindet
und die ganze Gegend durch eine besondere Größe der Pyramidenzellen
charakterisiert ist; die rückwärtige Partie der Area auf der vorderen
Zentralwindung ist sogar durch die bekannte Entwicklung von Betzschen
Riesenzellen vor dem ganzen übrigen Gehirn ausgezeichnet. Schon MEYNERT hat in
dieser ausschließlichen Entwicklung der Pyramidenzellen den Ausdruck für die
Motilität annehmen zu dürfen geglaubt, und BETZ ebenso speziell seit der
Entdeckung seiner Riesenzellen, die er als spezifisch motorische Elemente
ansah; die Klinik und das Experiment haben in großen Zügen dieser Annahme recht
gegeben. Die großen Pyramidenzellen der IIIc und V. Schicht, welche man wohl
für die Motilität in Anspruch nehmen kann, reichen jedoch über die zwei
agranulären Areae hinaus, und zwar über die schwach granuläre Area FC und den
rückwärtigen, auch schon granulären Teil von FD, den wir als FDm bezeichnen,
ferner über die ganze dritte Frontalwindung bis auf deren Orbitalteil, welche
überhaupt schon in ihrem Fusse granulär ist, also FBCm, FDΓ und FF. VOGTs
neue klassische Untersuchungen, in welchen er den architektonischen Bau der.
Rinde und den experimentellen elektrischen Rindenreizeffekt gleichzeitig
berücksichtigt, erlaubten ihm bestimmte Änderungen der motorischen Reizeffekte
zu konstatieren, welche mit gewissen Änderungen im Rindenzellbau parallel
gehen; eine nähere, recht eingehende Ausführung darüber geben wir später bei
Besprechung der Physiologie der einzelnen Areae (7. Kap., A, 3, § 7 und 10.
Kap., A, 5, § 7). Es scheint, daß man daraus sagen kann, daß die Area mit
Riesenpyramiden einen unmittelbaren Reizeffekt tonischer Spezialbewegungen
vermittelt, während die FB zwar vor allem neben etwas schwerer erregbaren
tonischen Spezialbewegungen auch sog. Einstellungsbewegungen (ganzer Komplexe)
vermittelt. Jedenfalls ist es an diesem Beispiele der Motilität interessant zu
sehen, daß einer anatomischen Änderung auch eine Qualitätsänderung im
motorischen Effekt entspricht, und jedenfalls scheint die Pyramidenrinde FA und
FB die spezifisch motorisch efferente Rinde zu sein. Daneben scheinen die große
Pyramidenzellen führenden Areae FC, FDm, FDΓ, evtl. auch FF, vielleicht
ebenfalls motorische Effekte neben anderen zu haben. Aber auch von der hinteren
Zentralwindung, Area PC, und vom oberen Parietallappen, Area PE, und auch
anderwärts sind motorische Reizeffekte zu erzielen. Jedenfalls ist aber eines
sicher: die Pathologie lehrt uns, daß die Motilität des Menschen vom Stirnhirn
derart abhängig ist, daß sie ohne dasselbe gar nicht möglich ist, während beim
Affen dieselbe sogar ohne Großhirn ohne weiteres möglich ist. Die progressive
Verrückung der Herrschaft der Zentren in der Nervenachse nach oben hat beim
Menschen ihr Maximum erreicht und zeigt den Sieg des Großhirns (JAKOBs) über
die ganze Motilität; dieser Sieg kommt in der kolossalen Entfaltung des
Stirnhirns zum Ausdruck in toto und auch im speziellen für die Motilität
einzelner bestimmter Partien. Man vergleiche deren Ausmaß (Area FA, B, C, D
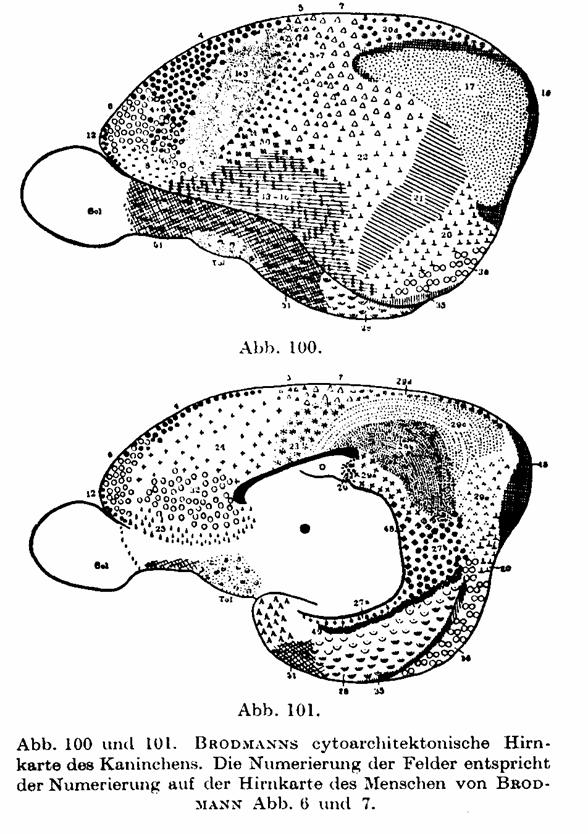
Abb. 92 oder Feld 4, 6, 8, 9 von BRODMANN Abb. 6) beim Menschen mit denen beim
Affen, Bär und Kaninchen (auf Abb. 100 und 101, 102 und 103, 104 und 105, 106
und 107), die aus BRODMANNs Lokalisationslehre entnommen sind. Ob man auf Grund
dieser Befunde den übrigen a granulären Areae FI, LA, LD, TGa, THa, evtl. auch
HE, motorische oder wenigstens efferente Funktionen zuschreiben kann, bleibe
vorderhand dahingestellt, vorstellbar wäre es, daß z. B. das Subiculum und
Ammonshorn, welches so exquisit aus Pyramidenzellen besteht, im Dienst des in
dem granulösen Koniocortex von HD lokalisierten Sinne (Geschmack? Geruch?)
efferente Impulse nach abwärts gibt, die nicht rein motorischer Natur sein
müssen, sondern mit der Salivation, Magensekretion usw. in Zusammenhang stehen
könnten, und FI und LA vielleicht solche in Abhängigkeit vom Geruchssinn
stehende efferente, evtl. sympathische Impulse (s. S. Kap., S. 450).
290 Das
Stirnhirn.
Daß aber auch die Betzschen
Riesenzellen in erster Linie in unmittelbarem Zusammenhange mit der motorischen
Funktion stehen, dafür sprechen, wie gesagt, auch manche pathologische Befunde.
So findet man bei amyotrophischer Lateralsklerose gleichzeitig mit der
Degeneration der Pyramidenbahn ein vollkommenes Schwinden der Betzschen
Riesenzellen aus der Hirnrinde (CAMPBELL, SCHRÖDER, ROUSSY und ROSSY). Doch
wissen wir aus eigener Erfahrung und aus CAMPBELLs Untersuchungen, daß hier
auch die übrigen Pyramidenzellen, ganz besonders die der IIIc-Schicht, und auch
die großen Pyramidenzellen der V. Schicht, ganz fehlen, und daß diese Atrophie
der Lamina ganglionaris sogar in das Gebiet der frontal davon gelegenen Area FB
übergreifen kann (SCHRÖDER, JAKOB, BUSCHER u. a. m.). Später greift die
Atrophie sogar auf die Zellen der IIIa- und IIIb-Schicht über. Ferner hat
CAMPBELL bei Leuten, denen vor langer Zeit eine Extremität amputiert worden
war, eine Veränderung der Betzschen Riesenzellen gefunden. Er fand dieselben
gequollen, fortsatzarm, sie hatten ihre Nisslschollen zum Teil verloren, der
Kern lag exzentrisch und war entrundet, eine Änderung, die man als Reaktion par
distance bezeichnen muß; ein eigentliches Zugrundegehen der Zellen, wie bei
amyotrophischer Lateralsklerose, fand er in diesen Fällen natürlich nicht.
Leider hat CAMPBELL, wie er selbst sagt, es unterlassen, auf die anderen Zellen
der Rinde bei solchen Amputationsfällen zu achten
Area praecentralis. 291
VOGT meint mit NISSL,
CHR. JAKOB (s. 1. Kap., S. 22), FOERSTER u. a. m., daß nur die V. und VI.
Schicht die Projektionsfaserung entsenden, während CAJAL, wie oben gesagt, und
wir daneben die großen Pyramidenzellen der IIIc- und auch der V. Schicht als Ursprungsorte
dieser Fasern ansehen, wenigstens betreffs der Pyramidenbahn. Erstere Ansicht
stützt sich außerdem noch auf BIELSCHOWSKYs, LENZ' und SPIELMEYERs
Untersuchungen von sog. "Lähmungen bei intakter Pyramidenbahn". Bei
dienen angeborenen Störungen fehlt die III. Rindenschicht beinahe vollkommen.
Diese Forscher nehmen nun an, daß die III. Schicht nicht der Ursprung der
Pyramidenbahn sein könne, da trotz des großen Ausfalles an Zellen, den die III.
Schicht in diesen Fallen bot, die Pyramidenbahn anatomisch intakt schien; sie
nehmen vielmehr an, daß hauptsächlich die Riesenzellen der V. Schicht die
Pyramidenbahnfasern entsenden müssen, da nie ebenso wie die Pyramidenbahn
selbst in diesen Fällen keine anatomische Veränderungen auf wiesen. Die
trotzdem bestehende Lähmung erklärten sie weiter aus dem Zellausfall der III.
Schicht dadurch, daß in der III. Schicht die Faseraufsplitterung der von außen
und speziell vom Thalamus in die Gehirnrinde einstrahlenden, die Erregung
zuleitenden Fasern stattfindet; die Zellen dieser III Schicht seien also
rezeptiv und fangen den Reiz dieser Erregung auf (s. S. 184); da nun bei ihrem
Ausfall die Aufnahme des Impulses zur Bewegung fehle, bleibe auch die Bewegung
als solche wirklich aus, und dadurch wird eine Lähmung gleichsam vorgetäuscht,
ebenso wie auch sonst motorische Lähmungen bei Durchschneidung sensibler Nerven
entstehen können. Es ist nun sehr möglich, daß diese Auffassung von
BIELSCHOWSKY und SPIELMEYER zu Recht besteht, ohne daß aus diesem Grunde
nötigerweise die Auffassung, die wir vertreten, daß auch die großen
Pyramidenzellen der IIIc- und V-Schicht Pyramidenfasern entsenden, damit in
Widerspruch stehen müsste; denn einerseits läßt sich bei einer im
Markscheidenbilde anscheinend "intakten" Pyramidenbahn nie erkennen,
ob nicht doch ein großer Teil der Fasern fehlt, deren Lücken durch die
vorhandenen verdeckt werden; andererseits könnte jener Teil der III. Schicht
doch noch zum Teil erhalten geblieben sein, der den Pyramidenbahnen ihren
Ursprung gibt, nämlich IIIc.
336 Das
Stirnhirn.
B. Regio frontalis (Tafel XIX—XXXI) (XLIV).
...
1. Area frontalis granularis FD
(Tafel XIX—XXX).
Die Formation erstreckt sich
ebenfalls von dem Sulcus callosomarginalis an der medianen Hirnwand über die
Mantelkante und die Konvexität des Gehirns bis in die Sylvische Grube und vorne
basal sogar teilweise bis an die Orbitalfläche des Stirnhirns, sie bildet also
im ganzen noch eine breite gürtelförmige Zone, die unmittelbar polarwärts von
der FC-Formation liegt. Der Stirnhirnpol selbst bleibt frei; er ist von einer
anderen eigenen Area FE eingenommen, die ihn kappenartig in der Größe ungefähr
eines Fünfmarkstückes überzieht. Abgesehen von diesem polaren Teile ist also
die ganze vordere Konvexität des Stirnhirns (vgl. Abb. 92) von der FD eingenommen;
ihre hintere Grenze fällt mit der vorderen Grenze der FC zusammen, reicht also
ebenfalls auf der ersten Frontalwindung weiter nach vorne, auf der zweiten
Frontalwindung springt sie nach hinten zurück und auf der dritten
Frontalwindung fällt sie zwischen Pars triangularis und Pars opercularis, also
ungefähr in den vertikalen Ast der Sylvischen Furche hinein. Die vordere Grenze
der FD an der Mantelkante ist dann ungefähr 5 cm weiter polar gelegen als die
hintere und zieht von hier in nach vorne konkavem Bogen um den Pol herum bis an
die Orbitalfläche. Auf diese Art bildet die Area frontalis granularis von
hinten nach vorne gerechnet das vierte Segment, welches das Stirnhirn in
frontaler Richtung halbringförmig umgibt (FA, FB, FC, FD) 1). [footnote p 336
1) Man vergleiche hierzu CHR. JAKOBs Sektorenbilder, die wir auf Abb. 17 und 18
wiedergeben.] Sie ist aber im frontocaudalen Durchmesser durchweg breiter als
die anderen Areae. Auch hier ist jedoch die frontocaudale Ausdehnung des
Gürtels dorsal an der Mantelkante größer und ventral in der Gegend der
Sylvischen Grube schmäler. Das in der Sylvischen Grube liegende ventrale Ende
dieses Ringes liegt auf der Pars triangularis und zieht auf dieser in die
Sylvische Grube hinein. Vorn auf der zweiten Frontalwindung sendet sie nach
vorn und unten an die Orbitalfläche gewöhnlich noch einen Fortsatz, der bis an
den Sulcus orbitalis lateralis heranreicht. In dieser weiten Ausdehnung zeigt
die FD verschiedene Abweichungen von ihrem Grundbau, zum Teil kleinere Modifikationen,
die hauptsächlich die Größe ihrer Zellen anlangen, oder Übergangsbildungen an
der Grenze gegen die benachbarten Areae, zum Teil wirkliche Varianten, die
einen konstanten, eigenartigen Bau aufzuweisen haben, wie FDL, FDΔ,
FDΓ.
770 Lobus
limbicus inferior.
...
Es würde noch die Frage
erübrigen, mit welcher Schicht der homotypischen, isogenetischen Formation wir
die Pyramidenschicht dieser allogenetischen, rudimentären Bildung
homologisieren dürfen. BRODMANN und nach ihm LANDAU haben die Ansicht
ausgesprochen, daß es die VI. Schicht sei, daß also alle übrigen Schichten beim
Übergang von der homo- und heterotypischen Rinde in die allogenetische
haltmachen und nur die I. und die VI. in die allogenetische Rinde übertreten.
Wir geben gerne zu, daß an manchen Stellen beim Menschen und wohl auch bei
Tieren es sehr den Anschein hat, als ob dies wirklich der Fall wäre und
verweisen hier auf die Tafel CV, wo der Übergang der drei Uncuswindungen
ineinander zu sehen ist. Auf der mittleren Windung 2 ist die Formatio der Area
uncinata, und zwar HA2, zu sehen mit der doppelt weißumränderten V. Schicht. In
der rechten Wand dieser Windung — bei Pfeil 2 bis Pfeil 3 — haben wir den
letzten vordersten Zipfel der Area granulosa HD, und es macht hier den
Eindruck, als ob jenseits von dieser Area nur ausschließlich die VI. Schicht
sich in schönem, nach oben konkavem Bogen um das Windungstal schlingen würden,
um darüber die Pyramidenschicht der Area pyramidalis unci auf den beiden hier
sichtbaren Gyri digitati derselben zu bilden, Kuppe 1 und 1'. (Nebenbei gesagt
sieht man hier auch einen gewissen lockeren Zusammenhang der Zellen dieser
Pyramidenschicht mit den Zellen des Nucleus Amygdalae (N. A. bei Höhe 24 /
Breite 28 cm ). Aber an anderen Stellen hat man oft einen ganz anderen
Eindruck, z. B. auf Tafel CX sieht man, wie die V. Schicht, die man aus HC
durch HD bis nach HE verfolgen kann, die ganze Pyramidenschicht der HE
gleichsam bildet, während die VI. Schicht eigentlich nur als ganz schmaler
Streifen darunter unter die Pyramidenschicht der HE zieht und diesen Saum bloß
bildet. den wir bei Besprechung der Area HE als Py (b) bezeichnet haben, und
der tatsächlich aus großenteils Spindelzellen und horizontalen Zellen besteht.
So ließe sich wohl bald das eine, bald das andere Beispiel für die eine und die
andere Ansicht anführen, und man muß es als wahrscheinlich bezeichnen, wie
schon auch aus der Zellform hervorgeht, daß sowohl die V. als die VI. Schicht
in die Pyramidenschicht des Subiculums hinein verfolgt worden können und mit
ihr direkt zusammenhängen. Phylogenetisch scheint auch die Ansicht, daß beide
Schichten der inneren Hauptschicht sich an der Bildung der Ammonswindung
beteiligen, besser begründet; man sehe diesbezüglich JAKOBs Genese der
Fundamentalschichten nach in Kapitel 1, S. 20—24, siehe .auch im folgenden § 6,
S. 787. Auch bei den übrigen allogenetischen Formationen, z. B. dem Übergang
der Area ultracingularis in die Area obtecta, haben wir auch die V. Schicht im
Zusammenhang mit diesen allogenetischen Bildungen gefunden (s. S. 470). Hier in
HE scheint die III. Schicht der HD nicht in Zusammenhang mit den Zellen der HE,
höchstens mit den Zellen ihrer großzelligen Glomeruli könnte ein Zusammenhang
(in HE1alpha) bestehen, Uns will es aber scheinen, daß aus solchen
"Zusammenhängen" an der Grenze zwischen Iso- und Allocortex überhaupt
lieber nicht auf eine Homologie der Schichten geschlossen werden sollte, denn
der Zusammenhang ist etwas Lokalanatomisches, und bei einer Entstehung aus
verschiedener "Anlage", wie es bei den allogenetischen Formationen,
besonders bei den rudimentären und primitiven, der Fall ist, ist jeder
Vergleich und jede Homologie von vornherein vergeblich und der anatomische
Zusammenhang ein trügerisches Hilfsmittel (siehe auch das 2. Kapitel: über die
Entwicklung der Rinde). § 5, 6 und 7 gemeinsam für die ganze Regio hippocampi
S. 771—792.
Area dentata. 787
Nach FLECHSIG gehört dieses
Gebiet des Uncus und des dorsalen Teiles des Gyrus hippocampi zu den
frühmarkreifen primordialen Sinneszentren, und er bezeichnet es als 4a und 4b.
Während er den Uncus zum Riechhirn rechnet, zählt er das Subiculum hippocampi
und den hinteren Teil des Gyrus fornicatus (LE) zur Geschmacksphäre! —
Physiologisch dürfte der Uncus infolge seiner unmittelbaren Verbindung mit dem
Tractus olfactorius nicht als Rinde zu bewerten sein, sondern wahrscheinlich
als Ganglion. —
Wir haben vorher schon erwähnt,
daß die Ammonsformation von einigen Forschern, darunter BRODMANN, als bloße
Fortsetzung der VI. Schicht angesehen wird (S. 771) und haben die Gründe, die
für und gegen eine solche Annahme sprechen, dort angeführt. Die
phylogenetischen Studien von ARIENS, KAPPERS, CHR. JAKOB u. a. haben zur
Erkenntnis geführt, daß das Ammonshorn mit seiner einen einzigen Zellschicht
keine Abortivrinde ist, sondern als eine primitive Rinde anzusehen ist; und
zwar als die allererste Rindenanlage (s. Abb. 15 und 16 a), welche sich aus dem
Corpus striatum in das bishin häutige Pallium vorschiebt und auf dieser
primitiven Stufe durch das ganze Tierreich und die ganze Lebensdauer hindurch
bleibt. KAPPERS nennt die Ammonsformation daher Archipallium, demgegenüber
nennt er die übrige Riechhirnrinde Paläocortex (Uncus usw.) zum Unterschied vom
Neocortex, welcher die ganze übrige Großhirnrinde ausmacht. (Wir haben schon
erwähnt, daß der Neocortex ungefähr unserem Isocortex [homotypisch und
heterotypisch] entspricht und der Paläocortex und Archicortex unserem
Allocortex.) Das Archipallium (Archicortex) hat bloß eine Staffel (V und VI =
innere Hauptschicht= innere Fundamentalschicht nach CHR. JAKOB), die beiden
anderen Rindenarten zwei Zellstaffeln, d. h. die eben genannte innere und dazu
die äußere Fundamentalschicht. Nun meint LANDAU, daß diese äußere Staffel im
Paläocortex (Uncus) eine andere sei als im Neocortex und daß sich auch an
Längsschnitten diese äußere Hauptschicht II und III des Uncus deutlich von II
und III des Isocortex durch die Färbung unterscheide und absetze. Betreffs der
Fortsetzung der Gebilde des Archipalliums auf das Retrosplenium kommt LANDAU zu
einer ähnlichen Auffassung, wie wir es auf Abb. 129 dargestellt haben; auch er
sieht die Taenia tecta und den Gyrus subcallosus als eine Fortsetzung der
Ammonszellschicht an (wie wir LF1 Area ultracingular. post, und LF2 Area
obtecta als Fortsetzung von HE1 und HE2 fanden). Ferner sagt er, die Zellen des
Gyrus intralimbicus seien die gleichen wie die der Fascia dentata, und er hat
insofern recht, als die Zellen im hinteren Induseum dieselbe Form haben wie die
Körner der Fascia dentata.

Professor von Economo

Croquis de este
sitio - Outline of this site
Índice - Table des matières - Inhaltsverzeichnis -
Table of Contents
Se recomienda mucho leer
o imprimir algunos de los trabajos mas extensos con el programa Acrobat Reader,
que puede obtenerse gratuitamente pulsando aqui 
Haga doble "click" en el título de cualquier
artículo, para leerlo ahora - Double-click on any article to read it now:
*
SOCIOLOGÍA DE LAS NEUROCIENCIAS
Pour comprendre l'enjeu : L’anthropologie
ganglionnaire, un psychovirus démasqué (français)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF
(190 kB: recomendada) o .DOC
(76 kB).
L'aliénisme en Argentine
: Diego Alcorta (1827) : Dissertation sur la manie... aiguë?
(français)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF
(600 kB: recomendada) o .DOC
(320 kB).
*
ELECTRONEUROBIOLOGÍA
Efectos relativísticos en
biofísica cerebral:
An ESSENTIAL preprint: Effects of relativistic
motions in the brain and their physiological relevance (To be published in Helmut
Wautischer, ed., Ontology of Consciousness: A Modern Synthesis)
(English) (Tema: funcionamiento del cerebro y psiquismo)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF (496
kB: recomendada) o .DOC (227 kB).
MBYKYHÁPE
GUARANÍME SUMARIO Y PÁRRAFOS INICIALES EN
CASTELLANO SUMÁRIO EM
PORTUGUÊS ABSTRAKTI SUOMEKSI
*
Diversificación
de recursos electroneurobiológicos en la evolución del sistema nervioso:
Anticipo de impreso: ¡Alma 'e reptil! Los
contenidos mentales de los reptiles y su procedencia filética (Introducción a una
próxima edición de Biomedicina de reptiles por Juan Carlos Troiano y colaboradores)
(Castellano with a VERY LONG English abstract) (Tema: origen del sistema nervioso)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF
(733 kB: recomendada) o .DOC (406
kB).
SUMÁRIO EM
LÍNGUA PORTUGUESA SUMARIO
CASTELLANO
Anticipo de impreso: Electroencefalograma y cerebro en reptiles (Fragmentos del Capítulo 2 del libro mencionado, Biomedicina de reptiles por Juan Carlos Troiano y colaboradores)
(Tema: el electroencefalograma de los reptiles invierte una relación básica en el electroencefalograma de los mamíferos: con frecuencia el de los reptiles pasa a presentar sincronías cuando están despiertos y a desincronizarse cuando están dormidos, mientras que en mamíferos es al revés. Así, esta sincronización del electroencefalograma reptil no concurre con la desconexión sensitiva del entorno, que encontramos en el sueño mamífero.) (Castellano)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF
(354 kB: recomendada) o .DOC (236
kB).
*
Cálculo de potenciales
dentro de las células
Calcule intensidades eléctricas y magnéticas
en cada compartimiento neuronal: The
nervous principle: active versus passive electric processes in neurons (Explains how to calculate electric
and magnetic field strengths inside different neuronal compartments) (LONG
FILE IN ENGLISH with Bulgarian, Russian and Spanish abstracts/TOCs)
Podrá
leer, imprimir o guardar en su disco duro esta investigación en versión .PDF
(2 Mb): recomendada) o .DOC (1,5 Mb). También como .html comprimido (compressed .HTML folder: 0,5 Mb) .ZIP.
ENGLISH ABSTRACT
AБСТРАКТ
НА БЪЛГАРСКИ
SUMARIO CASTELLANO
РЕЗЮМЕ НА
РОССИЙСКОМ
ЯЗЫКЕ
*
NOCIONES
GENERALES
Conceptos:
Noticia general -- ¿Qué es
electroneurobiología? -- La atmósfera intelectual (all in Spanish) -- Main Technical Ideas / Conceptos técnicos principales (English and Spanish) -- El descubrimiento de la Doppelrinde (German and Spanish)
Historia de las
experimentaciones:
1883: el primer mapeo con electricidad en
todo el mundo ¡durante ocho meses! de un cerebro humano
consciente, realizado en San Nicolás, Provincia de Buenos Aires (Spanish)
Table
of Contents (partial) of "Sensing: a new fundamental action of
nature" (English) -- Índices
Recepción de los aportes de Chr. Jakob en la neurobiología germana
__
The comments on Professor Christfried Jakob's contributions made in 'The Cytoarchitectonics of the Adult Human Cortex' by Professors Constantin, Baron von Economo, and Georg N. Koskinas, rendered in English by Prof. H. Lee Seldon (English)
You can download a .PDF
(673 kB: recommended) or .DOC (1.29
MB) file for printing this work.
__
Los comentarios de Economo y Koskinas sobre las contribuciones de Christofredo Jakob, traducidos por M. Szirko (Castellano: todavía no disponible)
__
Morphogenetic versus morphofunctional theory
-- Franz J. Irsigler's intervention in the Behavioral and Brain Sciences' discussion on the implications of the "initial brain" concept for brain evolution in Cetacea (1988) (English)
Puede obtener un archivo .PDF
(256 kB: recommended) o .DOC (106
kB) para imprimir este trabajo.
__
Recepción de los aportes de Chr. Jakob en la neurobiología y la sociedad argentina
Piva y Virasoro - Christofredo Jakob, neurobiólogo: científico en diálogo filosófico
(Spanish)
Puede obtener un archivo .PDF
(391 kB: recommended) o .DOC (466
kB) para imprimir este trabajo.
__
Kurowski - Animal óptico, el hombre: aportes de Christofredo Jakob incorporados a la "biopsicosociología" de Juan Cuatrecasas en la ciencia argentina
(Spanish)
Puede obtener un archivo .PDF
(384 kB: recommended) o .DOC (814
kB) para imprimir este trabajo.
__
Kurowski - Animal ótico, o homem: aportes de Christofredo Jakob incoporados à "biopsicosociología" de Juan Cuatrecasas na ciência argentina
(Portuguese)
Puede obtener un archivo .PDF
(385 kB: recommended) o .DOC (813
kB) para imprimir este trabajo.
*
Panorama evolutivo:
Table of
Contents of "Sensing: a new fundamental action of nature"
(English)
-- Humour... -- Comment l’ hylozoïsme scientifique contemporain aborde-t-il la sélection
naturelle du parenchyme neurocognitif? (français)
*
Vom Tierhirn zum Menschenhirn - Die Organisation der grauen Substanz des
menschlichen Zentralnervensystems (German and Spanish)
-- Christofredo Jakob: su vida y obra (Spanish)
*
FILOSOFÍA DE LA CIENCIA - CONCEPTO DE TIEMPO EN NEUROBIOFÍSICA

You can download a .PDF
(1 MB: recommended) or .DOC (571
kB) file for printing this work.
RESUMEN DIVULGATORIO Y PARA ESTUDIANTES EN CASTELLANO
Puede leer, imprimir o guardar en su disco duro el resumen en castellano
en versión .PDF (204 kB:
recomendada) o .DOC (96 kB).
*
“ANTAGONISMO ENTRE CIENCIAS DURAS Y HUMANIDADES BLANDAS”
Hubert Markl: Dementia dichotoma: Der Zweikulturenwahn - La ilusión
de las dos culturas (Spanish and German) -- Vista del Sur: una nota a la American Association for
the Advancement of Science, Washington (Presidenta Jane Lubchenco) sobre
prioridades científicas (English)
*
MALFORMACIONES
Y PAPEL DEL ÓRGANO CEREBRAL
Christofredo Jakob: “Los Monstruos
Anencéfalos” (Spanish)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF
(346 kB: recomendada) o .DOC (280 kB).
*
BIOÉTICA
¡Nuevo! Éthique de la Bio-Éthique (français)
Puede leer, imprimir o guardar en su disco
duro esta investigación en versión .PDF (323
kB: recomendada) o .DOC (161 kB).
*
EL PRESUNTO DUALISMO CUERPO - ALMA
¡Nuevo! Soigner et Guérir ? Le rapport
soma-psyché et l’archéologie du divorce culturel entre Médecine et Philosophie (français)
Puede leer, imprimir o guardar en su disco duro esta investigación
en versión .PDF (373 kB:
recomendada) o .DOC (201 kB).
SUMARIO EN CASTELLANO
*
PSICOANÁLISIS Y FACILITACION PSICOSOMÁTICA DE LA ENF. DE ALZHEIMER:
“CELOS: Olvido, homosexualidad, desmemoria” [Temas: Celos y olvido. Los celos de los muertos. Celos y homosexualidad. Los significados de ‘Untergang’. La ‘destrucción’ del complejo de Edipo. Una hipótesis acerca de la desmemoria (destrucción de la memoria). La enfermedad de Alzheimer.] (Spanish)
Puede leer, imprimir o guardar en su disco duro esta investigación
en versión .PDF (502 kB:
recomendada) o .DOC (110 kB).
*
PERICIAS JUDICIALES Y CASUÍSTICA:
Pericia médica en un caso de lisencefalia [Temas: Derecho de incapaces. Fundamento científico de la tutela jurídica de la adjudicación de recursos para enriquecimiento experiencial (calidad de vida) de oligofrénicos con retardo profundo del desarrollo intelectual. Acuerdo de juspositivismo y jusnaturalismo al respecto.] (Spanish)
Puede leer, imprimir o guardar en su disco duro esta investigación
en versión .PDF (300 kB:
recomendada) o .DOC (150 kB).
*
NUESTRA GENTE:
Reseña biográfica: Ramón Carrillo, el Gran Sanitarista
Argentino (Spanish)
*
Scrub The Web