Gobierno de la ciudad de Buenos Aires
Hospital Neuropsiquiátrico
"Dr. José Tiburcio Borda"
Laboratorio de Investigaciones Electroneurobiológicas
y
Revista
Electroneurobiología
ISSN: 0328-0446

A medio siglo de la muerte de Christofredo Jakob,
1956-2006:
Fuentes de la concepción biológica de la Doble Corteza
por
Diego Luis Outes

El profesor Dr. Diego Luis Outes sucedió a Braulio
Moyano en el liderazgo de nuestra tradición. Trabajó en este Laboratorio
durante cuarenta y tres años, desde 1943 hasta 1986, y actualmente (2006), a
veinte años de retirarse a su ciudad natal en el trópico argentino, Salta,
continúa desde allí sus investigaciones. Además de la jefatura del Laboratorio,
se desempeñó como catedrático titular de Anatomía y Fisiología del Sistema
Nervioso y profesor adjunto de Cínica Neurológica en la Universidad de Buenos
Aires y desde 1973 hasta 1982 fue director del Centro de Investigaciones
Neurobiológicas del Ministerio de Salud.
Contacto / correspondence:
Postmaster[-at]neurobiol.cyt.edu.ar
Electroneurobiología 2006; 14
(2), pp. 3-35; URL <http://electroneubio.secyt.gov.ar/index2.htm>
Copyright ©2006 Electroneurobiología. El presente es un
artículo de acceso público; la copia exacta y redistribución por cualquier medio
están permitidas bajo la condición de conservar esta noticia y la referencia
completa a su publicación actual incluyendo la URL original (ver arriba). /
This is an Open Access article: verbatim copying and redistribution of this
article are permitted in all media for any purpose, provided this notice is
preserved along with this article's full citation and original URL (above).
Imprimir este archivo no preserva la paginación
original. Puede obtener un archivo .PDF
(recomendado) o . DOC
para imprimir esta investigación, desde aquí o de / You can download a .PDF
(recommended) or . DOC file for printing, either
from here or
<http://electroneubio.secyt.gov.ar/index2.html
SUMMARY: Honoring Christfried Jakob's memory
in the fifttieth anniversary of his demise (1956-2006), the author examines one
of the most important concepts developed by the German-Argentine scientist: the
double cortex (Doppelrinde) idea. It
means, that all cortical regions possess a mixed functional character, so that
neither purely motor nor purely sensory areas are describable anywhere. By the way, it is also revisited the
notion that the deep cortical layers are motor and the superficial ones
sensitive, tracking down three forerunners of the idea: J. Luys, F. Nissl, C. U. Ariens Kappers.
SUMARIO: Con motivo de cumplirse
el quincuagésimo aniversario del fallecimiento del sabio germano-argentino
Christofredo Jakob, el autor revisa nuevamente una de las concepciones jakobianas
más trascendentes: la hipótesis de la "doble corteza" (Doppelrinde), según la cual todas las
regiones corticales son de funcionalidad mixta (son sensomotoras), no
existiendo cortezas motoras puras o sensoriales puras. Repasa, además, su idea
de que los estratos corticales profundos son motores y los superficiales
sensitivos o receptores, rastreando los precursores de esta concepción: J. Luys,
F. Nissl, C. U. Ariens Kappers.
1.
La contribución de Jakob
al Congreso Internacional
de Psicología de Munich
El 26 de noviembre de 1911, en el Congreso
Internacional de Psicología reunido en Munich, Christofredo Jakob pronunció una
conferencia. La disertación aparecería publicada el año siguiente (5) y se
titulaba "Sobre la existencia simultánea de una doble función sensomotriz
de la corteza cerebral como base de una nueva concepción biológica del órgano
psíquico cortical" ("Ueber die Ubiquität der senso-motorischen
Doppelfunktion der Hirnrinde als Grundlage einer neuen biologischen Auffassung
des kortikalen Seelenorgans").
Narraba ahí el resultado de sus estudios efectuados en
la Universidad de Buenos Aires (en este Laboratorio del entonces llamado Hospicio
de las Mercedes y en la Clínica Neurológica del Hospital San Roque, hoy
Hospital "Dr. Ramos Mejía") durante más de diez años y realizado en
base al estudio de focos cerebrales (más de 200, dice) como también
experimentalmente (Cebus del
Paraguay). Llegaba así a importantes concepciones neurobiológicas que podemos
resumir en lo siguiente:
A. - Todas las regiones de la corteza cerebral humana, y
también de los animales, eran activamente receptoras.
B. - De la misma forma no existía ninguna corteza que no
tuviera algo de motora. De ahí su apotegma: "En ninguna parte existe una corteza
que no sea al mismo tiempo receptora y efectora" ("Eine weder rezeptorische noch effektorische
Rinde (= 'Assoziationsrinde') existiert nirgends"). La corteza
cerebral era mixta en todas partes, lo único que variaba era la mayor o menor
proporción de lo sensitivo o de lo motor.
C. - Lo sensitivo y lo motor en la corteza presentaba cierta sistemática.
Eran receptoras las capas superiores (I a IV) y eran efectoras o motoras las
capas profundas (V y VI).
D. - Ambas capas corticales, desde los mamíferos con tejido
gris más indiferenciado hasta el hombre, tenían un doble origen, y lo mismo
sucedía en los vertebrados desde los reptiles hacia delante. (Jakob estudió también lo que, por entonces, estimaba era un taxón
de anfibios que se le aparecía como una verdadera transición hacia los
reptiles. Tratábase de los gimnofiones, donde el hecho podía demostrarse con
toda claridad y, si hubieran sido anfibios de verdad, por primera vez en la
serie filética. En realidad se trataba de un equívoco taxonómico, que se
comentará más adelante y ocasionó que inicialmente Jakob no trabajase con
anfibios sino con reptiles de convergente morfología externa; advertido y corregido
el equívoco, que Jakob, siguiendo la inspiración de su maestro von Strümpell, tomó
con muy buen humor y comentó siempre que pudo para evitar su propagación, el
hecho no dejó consecuencias neurobiológicas, aunque catorce años después todavía
lo repitieran von Economo y Koskinas.). Antes de entrar a estudiar sobre todo
estos dos últimos puntos conviene recordar algunos hechos históricos vinculados
a la estratificación cortical.
2. Encuadre
histórico de la contribución de Chr. Jakob
2.1. Meynert. Las capas celulares corticales no fueron
estudiadas y documentadas modernamente hasta la aparición del trabajo de
Meynert en 1867 (11). Meynert no dio interpretaciones sobre el funcionamiento
de las distintas capas, aunque observó que aquellas regiones donde predominaban
las células pequeñas o granos tenían un particular carácter
sensitivo-sensorial.
Theodor Meynert (1833-1898) 
Dice Meynert en 1869: "Como sitio de hallazgo de
tales agrupaciones granulares que replecionan la cuarta capa cortical, conocemos nosotros la retina, el
1. olfatorio y, además, las sustancias gelatinosas del asta posterior de la
médula y de la raíz trigeminal descendente, formaciones que, sin duda alguna,
se adecuan y sirven a la conducción sensorial". "Teniendo en cuenta
las regiones donde predominan estos hallazgos, los elementos granulares deben
considerarse como los portadores de la conducción sensorial" y, por si
esto fuera poco, agrega que "en el lóbulo occipital, en su cara interna...
existe una corteza de 8 capas" donde "la formación granular se repite
tres veces (capas 3, 5 y 7)... y aquí tenemos, nuevamente, la yuxtaposición de
lo sensorial con una masa de elementos granulares" (12) (pág. 20).
Respecto al asta de Ammón, donde sólo había células
piramidales motoras grandes, le atribuyó un carácter motor apoyándose también
mucho en que en la epilepsia (de manifestaciones motoras fundamentales) era
frecuente el hallazgo de lesiones del asta de Ammón.
Los estudios de Betz (1874) permitieron comprobar la
especial estratificación del área precrucial del perro y de otros animales y su
similar estructura en el hombre (área prerrolándica). Bevan Lewis y Clarke (1)
en 1878 lo volverían a confirmar.
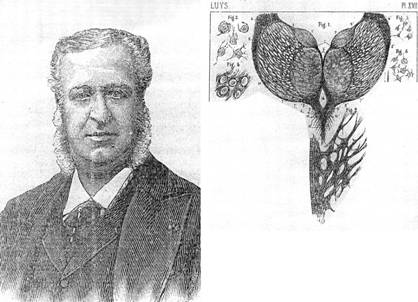
2.2. Luys. Pero por la época de Betz, y quizá
algunos años antes, la obra de J. Luys merece un párrafo aparte por la importancia
que guarda para nuestro tema. En realidad, y esto también lo acepta Brodmann, Luys
fue el primero en pensar en que cada capa cortical tenía una particular y específica función.
No era Luys un histólogo fino y eso lo detectamos en
su dibujo que reproducimos de su libro de 1876 donde no aparecen las capas granulares que tan bien
dibuja Meynert, si bien grafica entre ellas las células neuróglicas (fig. Nro.1). Pero era un pensador
profundo y claro. Veamos las palabras de su libro cuando habla de la significación
fisiológica de las regiones corticales: "... está permitido, apoyándose en
las leyes de la homología, suponer que las regiones submeníngeas ocupadas principalmente
por células pequeñas (son las c. piramidales pequeñas) deben estar sobre todo
en relación con los fenómenos de la sensibilidad, mientras que las regiones
profundas ocupadas por grupos de grandes células pueden considerarse como
regiones de emisión de los fenómenos de la motilidad. En efecto, aplicando a
este problema los datos adquiridos en el estudio de la médula espinal que
muestran, por ejemplo, que allá donde existen pequeñas células ocurren
fenómenos de la sensibilidad (astas posteriores) y que, al contrario, allá
donde existen grandes células son los fenómenos de la motilidad los que se
desarrollan (astas anteriores) es racional, digo yo, ver equivalentes
fisiológicos allá donde hay equivalentes morfológicos y considerar, como hemos
ensayado establecer, las regiones submeníngeas de pequeñas células de la
substancia cortical como la
esfera natural de la difusión de la sensibilidad general y especial y, por lo
tanto, el reservorio común de todas las sensibilidades reunidas del organismo.
Y, por otro lado, se pueden considerar las zonas profundas como los centros de
emisión y de preparación de las incitaciones a la motilidad. Esta manera de
considerar la corteza cerebral en su conjunto como un aparato esencialmente
sensitivo-motor concebido bajo el mismo plan
que los aparatos sensitivo-motores de la médula espinal, nos permitirá formular
algunas proposiciones nuevas" (10).

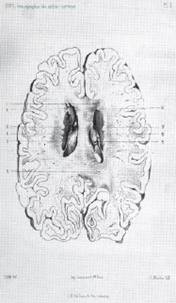
Fig. 1. (arriba): Cómo Jules
Bernard Luys (1828–1897) veía la
corteza cerebral. Redibujado de la fig. 1 del libro de Luys, Le
cerveau (1876). A, es la región "del retículo
submeníngeo de la neuroglia". B,
la zona submeningea de las pequeñas células piramidales, "región del
sensorium commune", dice. C, zona intermedia y D, la zona de las grandes células
piramidales. Nótese qué bien dibuja las células de la neuroglia, cosa que no
hace Meynert, quien junto con Luys es también de los primeros en graficar las
células corticales. Dibujo de Miguel Quiri (1975). Luys, un discípulo del microscopista clínico Charles Robin
(1821-1885), a sus 28 años escribió
e ilustró una celebrada monografía sobre el empleo del microscopio en la
anatomía patológica, diagnóstico y tratamiento de las enfermedades. Excelente
dibujante, daguerrotipista y luego fotógrafo, jefe de clínica en la Salpêtrière
y la Charité y luego director de manicomios, publicó Recherches sur le système cérébro-spinal, sa structure, ses fonctions
et ses maladies acompañado de un bello atlas cerebral tridimensional que él
mismo había dibujado a mano. Allí describió al núcleo rojo como "olive supérieure"; y como "bandelette accessoire des olives supérieures"
al núcleo subtalámico que Meynert en 1872 creyó una subdivisión de la
substancia nigra y en 1884 denominó discus
lentiformis y que, luego aún, Auguste Forel (1848–1931) llamó cuerpo de
Luys (corpus Luysii). Luys lo
estimaba un centro diencefálico para dispersar sobre el estriado la acción
cerebelosa, pero a través de sus conexiones con el pálido opera como inhibidor
del movimiento: su lesión determina hemibalismo, término éste asignado por von
Economo y separadamente por Kussmaul. Luys trazó asimismo la proyección desde
el cuerpo de Luys al globus pallidus y la proyección desde la corteza cerebral
al cuerpo de Luys, tractos ambos críticos para la fisiopatología del Parkinson
y objetivo mayor de la estimulación cerebral profunda. Luys creía que el corpus
striatum era el centro motor subcortical y el tálamo el sensorium commune, donde distinguía cuatro centros, cada uno
mediador de un sentido: el centro anterior u olfatorio, el centro medio u
óptico, el centro mediano o somatoestésico, y el centro posterior o acústico.
En sus últimos años, Luys se dedicó a la histeria y la hipnosis,
comprometiéndose en experimentos extravagantes como algunos destinados a
averiguar la acción a distancia de la medicación. Fue designado en la Academie de Médecine y recompensado con la Legion d'Honneur en 1877; fue fundador y primer director de L'Encéphale. Abajo: reproducciones, a la
albúmina (izq.), grabado (retrato), dibujo a mano y litografía de Luys. La
primera y la última imagen son de la Iconographie
photographique des centres nerveux, 1873; el retrato es de una nota
necrológica (1897) y el dibujo de sus Recherches
(1865).
No puede pedirse un pensamiento más claro. Empero,
Jakob no lo menciona. Por la época del trabajo de Jakob ya el método de Nissl,
descubierto en 1885, se imponía en todos los laboratorios europeos. En la
Argentina hay publicaciones que demuestran su precoz uso esporádico y hasta
ensayos con anilinas verdes, amarillas o anaranjadas: no sólo magenta, azules o
violetas. En Europa el Nissl se hizo,
con bastante rapidez, el proceder habitual para teñir células nerviosas. Prontamente
también fue utilizado para estudiar las diferentes características histológicas
de la corteza, siendo al respecto Carl Hammarberg el primero en realizar esto
en forma sistemática en 1895 (Studien über Klinik und Pathologie der
Idiotie nebst Untersuchungen über die normale Anatomie der Hirnrinde, Berlin
y Upsala – Nova Acta Reg. Soc. Sci.
Ser. III:1-126 Ak. Buchdruckerei Edv. Berling, traducción al sueco que fue el
deseo final del autor; un breve libro de 124 pp. con seis bellas litografías
desplegables), seguido por Brodmann (cuyo primer trabajo aparece en 1903) y
luego por Campbell (1905).

Korbinian Brodmann (1868-1918)
Surgieron entonces los mapas citotectónicos, que
cautivaron tanto a los neurólogos en una época en que el evolucionismo
familiarizaba a todos los investigadores con los estratos geológicos, y a los
que se agregarían, a partir del influjo de los Vogt, los estudios corticales
mielínicos, utilizando el método de Weigert o similares. Desde 1899 sería justamente
Jakob, bávaro como Nissl, el encargado de hacerlo conocer y propagar en la
Argentina.



Karl
Weigert (1845-1904), Oskar Vogt (1870-1959) y Cecile Mugnier-Vogt (1875-1962)
en fotografía y en estampilla postal. Abajo, juntos en su laboratorio.

2.3. Nissl. Descubierta la degeneración retrógrada por Nissl en 1894
tuvieron los investigadores en la mano un hermoso método para establecer el
origen de las fibras nerviosas en las distintas agrupaciones nucleares. Es muy
del sabio germano-argentino no mencionar fichas bibliográficas, costumbre que
hace difícil determinar de donde arrancan sus ideas. Tal lo que me ha sucedido
en este trabajo. Me cuesta buscar el hilo conductriz.


Franz Nissl (1960-1919). El retrato de la izquerda fue
confeccionado en 1981 por Francisco Pérez Pergaux,
colaborador por muchos años del autor e ilustrador de un buen número de mis artículos
sobre temas históricos. Pergaux había
estudiado con Salvador Dalí y la dolencia que lo hizo paciente crónico del
Hospital finalmente afectó un solo aspecto de sus retratos: la mirada, que
perdió su lograda expresividad. Pergaux
falleció después que yo me jubilase en el hospital. Ello habrá ocurrido unos
diez años después de capturar la actitud de Franz Nissl en este excelente
retrato.
Yendo a las relaciones tálamo-corticales – en que se
basa en buena parte el trabajo de Jakob, como luego veremos – el primero en
estudiarlas fue Gudden (1876) con su método de extirpación de regiones
corticales de animales jóvenes y posterior observación de la degeneración
nuclear (celular y mielínica) correspondiente. Para ello desarrolló también
mejores micrótomos junto a Auguste Forel. Este método dio un fuerte argumento a Forel (1886)
a favor de la teoría neuronal que
empezaría a fundar junto con His (1887)
y Cajal (1889).


Auguste-Henri Forel (1848-1931), mirmecólogo,
neuroanatomista, psiquiatra y reformador social suizo activo con Magnus
Hirschfeld y Havelock Ellis en la Weltliga
für Sexualreform (que calificó la
doctrina racial, "die Eugenik",
como "die Kakogenik") y fue
influyente sobre Cesare Lombroso, Sigmund Freud, Alfred Adler, Wilhelm Stekel y
Paolo Mantegazza. Obtuvo notables resultados en sus investigaciones de la
estructura del órgano cerebral humano y del ganglio cerebral de las hormigas.
Su amigo el eminente entomólogo Horace Donisthorpe se quejó de que el tratado
sobre las hormigas permitiera a este reformador de prisiones y asilos
"ventilar sus ideas socialistas". Irónicamente la banca suiza puso su
efigie en uno de los billetes de mayor valor (derecha)
El método de Gudden alcanzaría su máxima victoria
neuroanatómica al ser utilizado por C. v. Monakow en su célebre y monumental
trabajo sobre las relaciones tálamo-corticales de 1895.



Bernhard
von Gudden (1824-1886, izquierda), catedrático de psiquiatría en Munich, médico
del rey Ludwig II de Baviera, con él murió ahogado en Pentecostés de 1886, en
el lago Starnberg. Eduard Hitzig (1838-1907, centro) y su asistente Constantin
von Monakow (1853-1930).
Estas relaciones tálamo-corticales fueron estudiadas
con el método de la degeneración retrógrada por primera vez por Nissl, sobre
cuya obra nos detendremos por que tiene el gran mérito de haberse puesto a
pensar, después de Luys, en el significado de cada capa cortical en si misma: no tanto en el del
conjunto de las capas. Si se me permite la expresión, la tendencia exploratoria
de Nissl era más "horizontal", la tendencia de los hombres de la
época más vertical. Por eso Nissl no se ocupa de la citotectónica a pesar de
ser el descubridor del método ideal para ello. Algo que sorprende a cualquier
historiador. Como si para él fuese algo de poca importancia.
Según Spatz (19) ya Nissl había advertido en su
trabajo de 1907 que "las alteraciones patológicas corticales, hoy demostrables
ya no raramente, predominan en determinadas capas corticales" (16). En
pocas partes pudo encontrar alguna relación entre estos hechos y la
distribución de los vasos sanguíneos.
Afirma Spatz que Nissl ya en sus tiempos en Frankfurt (1895-1904) había tratado de
investigar experimentalmente las relaciones tálamo-corticales, pero que recién
en 1907 comunicó sus primeros resultados. Refiere que pudo separar
completamente en un perro recién nacido la corteza de los núcleos basales. Al
sacrificar el animal, a los meses, observó lo siguiente:
1. - La corteza aislada del perro recién nacido continuaba su
desarrollo a pesar de su aislamiento y a pesar de no haber funcionado nunca en
su debido marco fisiológico.
2. - A pesar de que toda la corteza estaba desprovista de
influjos externos y de que se diferenciaba
por si misma, todas las capas no se comportaban igual. Las inferiores (V y VI)
mostraban (con su método por supuesto) una severa decadencia celular, mientras
que en las capas superiores esta tendencia era mucho más escasa. De esto infirió
Nissl que las diferentes capas corticales estaban unidas en distinta forma a
los órganos centrales (tálamo sobre todo): las dos capas profundas mantendrían
una mayor unión y las capas superficiales, que poco sufrían en la preparación,
tenían una relación más floja, una mayor independencia, y esto sucedía en casi
toda la corteza independientemente de la división en áreas.
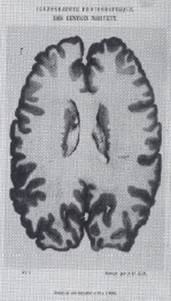
En la figura adjunta reproducimos la imagen con que
documentó Nissl este hallazgo (fig. 2).
Aunque Spatz no dice donde apareció la microfotografía de Nissl, he tenido la fortuna
de encontrarla en un trabajo de 1911 (17).
A la luz de los conocimientos actuales sabemos que lo
que veía Nissl era exacto y que su documento es irrefutable, pero la
interpretación no es correcta. Al aislar la corteza quedan cortadas tanto las
vías tálamocorticales como las corticotalámicas y corticocorticales: la
destrucción de estas vías eferentes provoca la degeneración retrógrada y
posterior destrucción de las células motoras de la corteza y si ello ocurre en
las capas V y VI es porque justamente ahí están acantonadas las células
motoras. En cambio el corte de las vías tálamocorticales provoca degeneración
retrógrada en el tálamo y nada en la corteza, puesto que el método de Nissl no
hace visible la destrucción del axón separado de su célula ni los botones
terminales. Lo que Nissl veía era la demostración de que las capas inferiores
eran fuertemente efectoras, siendo un error considerar que la unión de las
capas superficiales con el tálamo era menos estrecha que la unión de las
inferiores.


Fig. 2. -Microfotografía de Nissl que publica Spatz en su
trabajo de 1929 (apareció publicada en un trabajo de Nissl de 1911); pertenece
a un perro. La corteza inferior es normal (der.). La superior (izq.), en la
misma zona del perro pero después de "aislar" la corteza. De acuerdo
a esta demostración, Nissl consideró que las capas superiores de la corteza
"estaban en alto grado en relación directa y exclusiva con los fenómenos
psíquicos". Confirmaba los atisbos geniales de Luys.
Nissl no extrajo otras consecuencias fisiológicas de
sus experimentos aunque dice Spatz (19) que en la pág. 73 de su Neuronenlehre (15) estas consecuencias
surgen casi forzosamente. Hemos ido en busca del complicado libro de Nissl (uno
de los libros más engorrosos que nos ha tocado leer y donde el autor alemán
expone sus ideas sobre su "nervoses Grau" y donde se declara
reticularista) para aclarar ese párrafo de Spatz y hemos hallado las siguientes
ideas de Nissl:
En primer lugar separa tres zonas en la substancia
gris; a) la corteza blanca (Rindenweiss),
b) la corteza gris (Rindengrau) y c) la corteza del techo (Rindendach).
La corteza del techo constituye la capa superior de la
corteza, la zona libre de células, tan bien delimitada. Con toda seguridad,
dice Nissl, que esta capa o techo cortical es el de menor valor fisiológico. Y afirma que esto ya lo
decía Meynert en 1872, al expresar que esta capa no podía tener "gran
dignidad neurológica".
La segunda zona, la corteza gris, comienza en el límite
inferior de la anterior y engloba la capa de las pequeñas y grandes piramidales y también, en ciertas partes, la
capa granular o 4ta.
de Meynert. La tercera zona es el resto de la corteza formada por una capa
interna de células fusiformes y otra externa de células piramidales grandes
(son la V y la VI actuales), es la zona de las fibras mielínicas radiales. Y
luego agrega textualmente Nissl: "La corteza blanca es, en cierto modo, la
antecámara de la corteza gris, constituye la puerta de la corteza, tanto la
puerta de salida como la de entrada y también la puerta lateral hacia las
diferentes regiones corticales". La puerta lateral de Nissl es la región
por donde salen y entran las fibras asociativas cortas (fibras en U) y largas.
Eliminando la primera zona cortical se pregunta luego Nissl cuál de
las otras dos zonas tiene mayor valor funcional y junto a la palabra funcional coloca su pie de página número
4, al que se refiere Spatz y que dice: "La expresión funcional se refiere aquí, sin excepción, a las funciones psíquicas
de alto valor fisiológico, significa aquí un tejido que se adecúa directa y
exclusivamente para los hechos psíquicos, como otro de menor valor o de
funciones más primitivas significa un tejido cuya acción nada tiene que hacer
con los fenómenos psíquicos o bien que no está limitado exclusivamente a ellos
realizando también otras funciones nerviosas."
¿Qué relación tienen estas ideas de Nissl con el
experimento en perros de 1907-1908? Pues, sencillamente, que en los animales
también las capas superiores por estar más aisladas de los núcleos basales, por
tener menos relación con las eferencias, debían servir para elaboraciones de
más alto nivel. Este experimento, aunque no pudo comprobarlo patológicamente en
el hombre, lo ayudó a pensar que en éste la zona gris o media debía ser la más
importante, máxime cuando la zona inferior o blanca estaba casi repleta por las
fibras radiantes de la corteza y casi desaparecía en la profundidad de los
valles cisurales.
La expresión
"portal de la corteza" es feliz pues es exacto que las capas
inferiores emiten sus eferencias a través de este portal y que por él entran
las aferencias. Pero decir que las capas inferiores son la "antesala"
de las capas superiores no lo es tanto y es dejar de lado los descubrimientos
de Cajal (18), según los cuales la corriente aferente llega directamente hasta
el estrato superior y que este, en verdad, es un estrato sensitivo psíquico.
Fue Cajal en sus estudios corticales de 1899 el primero que demostró esta
llegada de los impulsos aferentes hasta las capas superficiales de la corteza y
Nissl, tenaz enemigo de la teoría neuronal, no se refirió a ello.
En resumen, Nissl, después de Luys, en base a hechos
experimentales, fue el primero en pensar en la existencia de dos estratos
paralelos y superpuestos en la corteza cerebral y atribuir distintas funciones
a ambos: el estrato inferior sería motor y receptor al mismo tiempo y el
superior el estrato psíquico fundamental. Volvamos ahora al trabajo de Jakob de
1911-1912.
Fundamentos
de la impugnación de Jakob a Flechsig
¿En que se basaba Jakob para afirmar que toda la
corteza era activamente receptora? No debe olvidarse que esta afirmación era
levantarse nada menos que contra Paul Flechsig. Este, en base a sus estudios
mielogenéticos, había llegado a la conclusión de que los grandes centros
asociativos de la corteza no proyectaban fibras: ni recibían ni emitían.
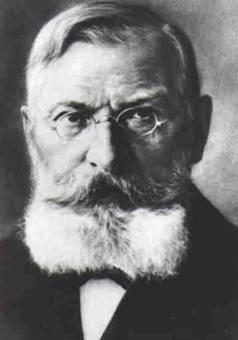

Paul Emil Flechsig (1847 - 1929) y su discurso
inaugural al asumir el rectorado (este ahora está traducido al castellano por el
presente autor)
Jakob para fundar su teoría tuvo que basarse en la
existencia de las relaciones tálamocorticales, estudiando, en focos corticales,
la degeneración retrógrada talámica.
Con el método de Nissl-Lenhossek encontró así que los
focos frontales (a excepción de los orbitarios) provocaban lesiones en el
núcleo lateral anterior del tálamo; los focos del girus supracalloso
repercutían en el núcleo anterior y dorsal del tálamo; los del precuneus y
parietal posterior en el pulvinar anterior y dorsal; los del temporal posterior
en la base del pulvinar y los del temporal anterior en el núcleo basal interno del tálamo. En cuanto a la relación del centro
acústico y del óptico con los geniculados, que tan bien había sido puesta de
manifiesto por Monakow y Flechsig, no era necesario ni discutirlo.
Jakob llegó a la conclusión de que para cada forma de
la sensibilidad existían dos territorios corticales vecinos: uno, que era el
centro proyectivo clásico o territorio primario directo de la corteza, unido al
tálamo basal, y el otro, que eran los
centros de asociación o territorios secundarios indirectos y que estaban unidos
al tálamo dorsal. Todas las vías de la sensibilidad que llegaban al tálamo basal
estaban unidas por vías de asociación o intratalámicas a los núcleos dorsales.
Toda la corteza entonces, directa o indirectamente, estaba vinculada a la
sensibilidad: era receptoramente activa.
La idea de Jakob de que las capas superiores eran
receptoras y las inferiores motoras, estaba abonada por varios hechos. Unos
eran de carácter exclusivamente histológico, pues en aquellos lugares donde
terminaban las vías sensoriales (centros proyectivos de Flechsig ) adquirían
gran desarrollo las capas superiores y casi desaparecían las inferiores y,
viceversa, en los centros motores por excelencia (corteza prerrolándica)
predominaban estas capas inferiores y disminuían en grado sumo las superiores.
La utilización del método de Weigert para demostrar la
disminución o desaparición de las corrientes aferentes corticales era muy engorrosa
y sólo podía realizarse en ciertos sitios, por ejemplo, la desaparición de la
estría de Gennari en caso de focos talámicos ventroposteriores. Lo mismo pasaba
con el estudio al Marchi. En realidad, el que demostró en forma más clara y
correcta la llegada de los impulsos talamosensoriales a las capas superficiales
de la corteza fue, repetimos, Ramón y Cajal con sus impregnaciones argénticas
(Golgi).


Camillo Golgi (1843-1926) y
Santiago Ramon y Cajal (1852-1934)
Jakob compara en su artículo lo que sucede en la
corteza con lo que ocurre en las estructuras nerviosas más simples: "así
como hay en ellas, en toda su extensión, columnas motoras y sensitivas, tanto
en la m. espinal como bulbo y cerebro intermedio, así sucede también en el
manto cortical donde por todas partes
ambas capas fundamentales corren paralelamente y, así como allá, de acuerdo a
los requerimientos periféricos, puede crecer una u otra columna, acá también se
cumplen leyes homólogas." " En la traducción que hizo Jakob de este
artículo en la Prensa Médica Argentina
(1916) agregó el siguiente párrafo: "Para la orientación futura atribuimos
conceptualmente a tal disposición sensomotora el mismo valor que tuvo el
descubrimiento de Bell y
Magendie respecto a la distinta naturaleza funcional de las raíces anteriores y
posteriores". Obsérvese el parecido de esta frase de Jakob con la que
transcribimos anteriormente de Luys.
Pero "el conjunto del manto cortical actúa ya sea sensomotora o
motosensorialmente según el predominio de uno u otro componente y una parte de
la corteza que funcione y actúe solo receptoramente o solo efectoramente no es
demostrable ni histológica ni experimentalmente". "En ninguna parte
hay un tipo cortical que
merezca exclusivamente el nombre de corteza de asociación o de centros
conmemorativos". "Estos procesos son funciones de ambas capas
fundamentales en todas las regiones corticales y no son de naturaleza
localizatoria sino dinámica. En contra de las opiniones de Flechsig y Ramón y
Cajal debemos, entonces, reconocer, en principio, el valor equivalente de todas
las zonas corticales". "Todos los actos corticales son, por
consiguiente, y a priori, de naturaleza mixta, senso-motora, siendo la
separación de ambos componentes inconciliable con la textura cortical".
Para el entendimiento de la función cortical, aquel hecho era fundamental para
Jakob. Cada proceso elemental de la esfera volitiva o de la sensibilidad tenía
que llevar por eso, desde el principio, el mismo caracter mixto y era falso
hablar simplemente de procesos de la voluntad o de la sensibilidad; en
realidad, en cada caso predomina un componente sobre el otro: ambos son
diferentes no en su esencia sino en su tendencia. Por eso llegaba Jakob a un
concepto monístico de todas las funciones corticales, que cuando sus discípulos
pudieron fundamentar objetivamente la eficacia del psiquismo dejó claro para
estos que esa intimidad personal no podía ser ni solamente sensitiva ni
solamente volitiva: si su entorno cortical mostraba ambos elementos, es porque
ellos integraban la fuente eficaz incrustada en el arco sensoriomotor.
Esto mismo, todavía desde el reactivismo, es lo quería
significar Luys en 1875 con sus palabras: "Es así que uno llega a admitir
que desde el punto de vista fisiológico el acto motor voluntario que emana del
cerebro no es más que la repercusión más o menos alejada de una impresión
primordial primitiva", y en estas otras: "...así podemos decir que si
el acto de la voluntad no es más que un fenómeno de sensibilidad transformada,
es, sin embargo, la sensibilidad multiplicada por todas las actividades
cerebrales emocionales, en una palabra la personalidad vibrante y que entra en
juego en forma somática y que se exterioriza por una serie de manifestaciones
reflejadas y coordinadas". O bien esta última y contundente frase:
"La motricidad no es pues, fisiológicamente, más que la sensibilidad
transformada".
Anatomía
comparada de la corteza cerebral y su vinculación con el problema de la doble
corteza
Nada mejor para comprobar la diferente funcionalidad
de las capas corticales que estudiar la evolución filética de la corteza. Al
respecto, antes de aparecer los trabajos filogenéticos de Jakob (1911) ya Ariens
Kappers, en 1909, defendió la división cortical en dos capas, la superior de función receptiva-psíquica y la
inferior o efectora-motora.


Dos
imágenes del Prof. Kappers, cuyos tres nombres eran Cornelius Ubbo Ariens
(conocido también como si fuera apellidado Ariëns Kappers o Ariens Kappers,
1877-1946)
Ariens Kappers se basó en lo siguiente:
a) en los reptiles, en su arquicortex, halló un gran
predominio de las células granulares, casi dos veces el número de las c.
piramidales.
b) en los mamíferos inferiores ya se nota en el asta de Ammón
(arquicortex) un gran aumento de las células piramidales, adquiriendo la zona
funciones proyectivas y asociativas.
c) la formación en el asta de Ammón de un campo olfato-psíquico
(zona subicular) determina en los mamíferos la aparición de gran cantidad de
células piramidales por arriba de la capa granulosa inferior: son las células
piramidales supragranulares. Además de estas particularidades onto- y filogenéticas, cita Ariens Kappers hechos
patológicos, como los de un trabajo de Bing (1907). Bing, en un sordo (no dice
si central), encontró muy disminuídas (casi en sus 3/4 partes) a las capas
corticales inferiores de la corteza auditiva, habiendo hecho v. Monakow la
misma observación en un sordo con lesiones en la cápsula interna. Por último, Ariens
Kappers refiere que Winckler le mostró un caso parecido. De ahí que diga:
"las capas subgranulares dependen, en primer lugar, del desarrollo local
del sistema subcortical al cual pertenece dicha región, mientras que las capas
supragranulares dependen, principalmente, de las asociaciones interregionales
con las zonas corticales vecinas y distantes" (pág. 168).
Este párrafo es como una repetición de lo que
encontrara experimentalmente Nissl en 1908 (el trabajo de Bing es del año 1907,
el de v. Monakow de 1905 y el de Winckler es una comunicación personal de Ariens
Kappers). Las pirámides supragranulares son las últimas en aparecer
ontogenéticamente (como lo demostrara Mott en 1907) (13) y tendrían funciones
asociativas de elevado orden, según A. Kappers. Nótese que esta idea se hará
notar en Kleist, para quien el gran desarrollo de la tercera capa de células
piramidales pequeñas y medianas es propio de los centros psíquicos.
Jakob tiene el gran mérito de haber estudiado, con
particular atención, el encéfalo de las anfisbenas (víbora ciega) y, superado
el antes mencionado equívoco, los gimnofiones. En su trabajo de 1918 (7)
confiesa que ya había empezado en 1905 a estudiar el cerebro de los batracios y
reptiles argentinos. Las primeras comunicaciones, aunque parciales, las hizo en
su trabajo "La célula cortical en la
locura" (2), luego en trabajos de 1911 en Europa (3) y, posteriormente, ya
de vuelta en Bs. As. en
1913, en su Atlas de los mamíferos
argentinos (6). El clásico esquema de Jakob y que figura en todas sus
publicaciones es el que reproducimos (fig. Nro. 3) y reproducen siempre sus estudiosos, como por ejemplo von
Economo y Koskinas. Se observa en este dibujo, con toda claridad, la unión
del estrato externo (S E) con la corteza olfatoria (R H) y la del estrato
interno (S I) con el cuerpo estriado (C. ST). El esquema B pertenece a la comadreja (Didelphis azarae), donde también se repite esta imagen, ya en forma
más clara.
En el trabajo original de Jakob, en su pág. 37, dice
que los estratos corticales externo e interno "derivan" de la corteza
olfatoria y del cuerpo estriado respectivamente (3). En ninguna parte habla
Jakob de "migración" de estas células, aunque la utilización de la
palabra "derivar" lógicamente haga pensar que ese haya sido su
pensamiento.
Como recién se
anticipó, en todos sus trabajos iniciales (1911-1913) Jakob afirmaba que el
esquema donde tan claro se veía esta relación de los estratos corticales
pertenecía a un animal de la familia de los gimnofiones (Coecilia), animal que sería intermediario entre anfibios y
reptiles. Posteriormente, en numerosas conferencias y publicaciones como por
ejemplo la de 1918 (7), rectificó esa clasificación, reconociendo su error en
la manera en que – para la materia clínica – lo requería uno de los más
conocidos adagios de su maestro y amigo von Strümpell (una de las exigencias que a éste más se le
atribuye repetir era, "Jeder
vernünftige Arzt wird sich nicht für unfehlbar halten und wird einen Irrtum in
der Diagnose stets freimütig bekennen", que significa "Ningún
médico razonable se considerará infalible y reconocerá de inmediato y
francamente todo error de diagnóstico") y hallarse en presencia
de un animal del suborden de las anfisbenas (víbora ciega), más precisamente de
la Amphisbaena darwini. En este
trabajo de 1918 presenta dos cortes transversales, de un gimnofión y de una anfisbena
(animales que se parecen bastante en su morfología externa y comparten algunos
hábitats), donde se ve claramente que la anfisbena es un reptil, con su clásica
epidermis escamosa y con su cerebro mucho más desarrollado, mientras el cerebro
del gimnofión es vesiculoso, como el de un urodelo.
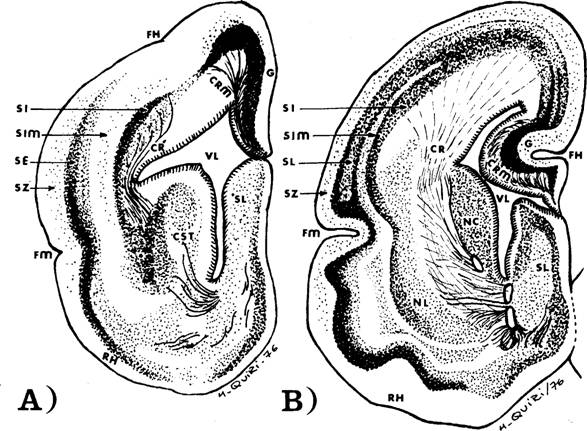
Fig. 3. - El esquema de Chr. Jakob que aparece, por primera
vez, en el Vom Tierhirn zum Menschenhirn y
en el Menschenhirn, ambos de 1911,
aquí redibujados por el voluntarioso agente administrativo de este Hospital,
Miguel Quiri; a) es el esquema de la
Amphisbaena darwini (vibora ciega)
que clasificó, por ese entonces, dentro del género de los gimnofiones,
rectificando luego el error; b) el
esquema de la comadreja Didelphis azarae:
si, estrato interno; se, estrato externo; sim, estrato intermediario; sz,
estrato zonal; cst, cuerpo estriado; sl, septum lucidum; erl, corona radiada lateral; Rh, rinencéfalo; fh,
fisura hipocampica; fm, fisura medial; ci, cápsula interna; g, fascia dentada; rb, radiación basal.
Cuando mi querido maestro Moyano (14) habla de la
concepción filética de su maestro, expresa: "Estudiando el cerebro de los
reptiles (la anfisbena darwini) pudo
comprobar que la parte externa de la corteza tiene su origen en la región
olfatoria, receptora, sensitiva y la parte interna en el cuerpo estriado,
núcleo efector, motor. Viejo conocedor de la historia filética de la corteza,
no tardó en aplicar esas observaciones a la organización fundamental de la
corteza humana".
Antes de aceptar la teoría filética de Jakob
deberíamos preguntarnos lo siguiente (ver microfotografía de la fig. 4): ¿desde el receso lateral del ventrículo (que marca
con una x) existe o no proliferación ependimaria y luego la posterior
migración? Nadie hasta ahora, que sepamos, ha demostrado que no y, entonces,
esta migración formará el extremo inferior del estrato interno: justamente la
zona que une este estrato con el cuerpo estriado. Esto hará pensar,
lógicamente, que este segmento puede derivar del estriado.
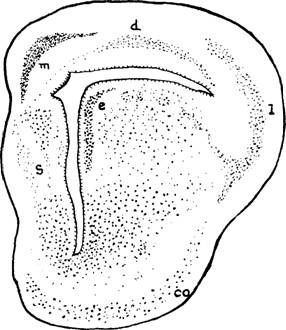
Fig. 4. -
El esquema de la Lacerta, en corte
oral al foramen de Monro, tal como lo grafica Kuhlenbeck en 1922. e, epiestriatum;
s. septum lucidum; b, núcleo basal; co, corteza olfatoria; I, área lateral
(neocórtex); d, área dorsal (archicórtex): m, área medial (archicórtex). Se
observa que el neocórtex no aparece biestratificado y que se prolonga abajo con
la zona del estriado. La zona dorsal (d) es la futura Asta de Ammón y la medial
(m) la futura fascia dentada. El extremo lateral del área dorsal parece
colocarse abajo del neocórtex en la posición que ocuparía el estrato interno.
Para mi manera de ver es más lógico pensar que el
estrato celular interno es el último "empuje" migratorio en dirección
a la pía, coincidiendo ello con la masa del estriado en el sentido de que éste
también es el último "empuje" de un sector más inferior de la
vesícula telencefálica. Todo ello hablaría de cierto parentesco entre estos dos
sectores celulares; ambos estarían originados por una misma "poussée" migratoria secundaria.
Kuhlenbeck (9), que seguía los principios
neurobiotáxicos de Ariens Kappers, ha escrito también un concienzudo trabajo
sobre la evolución filogenética de la corteza. Coincide a grandes rasgos con el
de Kappers y de allí extraemos el esquema de la Lacerta, que también redibujamos
(fig. Nro. 4, obra
de Miguel Quiri). En él notamos que el neocórtex o corteza lateral (1) hacia
abajo contacta con la zona del cuerpo estriado (epíestriatum) dando la
impresión de continuarse directamente. No hay biestratificación del córtex lateral. Es evidente que la biestratificación del córtex lateral o neocórtex primitivo es más marcada en la
anfisbena, como lo revela la microfotografía de Jakob que aparece en su trabajo
de 1918 (fig. Nro. 5).
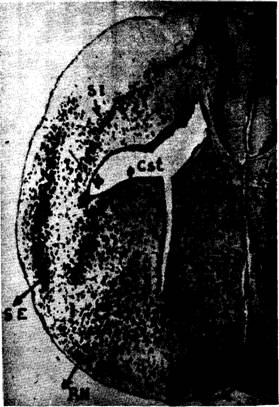
Fig. 5. -
La microfotografía de Chr. Jakob de la anfisbena darwini. Apareció en su trabajo de 1918. La reproducimos a mayor
aumento. Se vé el estrato interno (si) de la corteza lateral que se prolonga
hacia arriba hasta el asta de Ammón y hacia abajo hasta la región del estriado
(Cst). Jakob le dio más importancia a esta unión inferior que a la superior con
el asta de Ammón cuya función motora es tan indudable como la del estriado. En
la microfotografía se ve claramente que es más nítida la unión con la zona
amónica que con la estriada. s.l.; septum lucidum. Rh., rinencéfalo.
Esa es la importancia de la anfisbena para el estudio
del cortex primitivo. Por ello es
tan necesario que su estudio sea retomado por los investigadores para aclarar
hechos que en la realidad no aparecen tan esquemáticos como en los dibujos.
Kuhlenbeck muestra sólo dibujos, esquemas. Jakob presenta microfotografías.
Kuhlenbeck cita a Jakob en la bibliografía pero no en el texto.
Jakob da una prueba más de la duplicidad originaria
ontogenética de la corteza cerebral humana: "Al comienzo del 5º mes se
desarrolla una fase pasajera de la organización cortical y que, a nuestro entender, nadie lo ha constatado
todavía y reconocido en su fundamental importancia para la organización de la
corteza. Como se ve en la fig. 39 se
notan acá los elementos corticales dispuestos en dos capas paralelas. .. "
"Este tipo fetal biestriado corresponde completamente a la construcción en
dos capas fundamentales de la corteza, una conducta que no es sólo del hombre
sino que también se produce en toda la serie de los animales y es la expresión
del viejo origen de las elevadas y diferenciadas cortezas" (4).
Algunas de estas ideas ya las he expresado hace más de
treinta años, exponiéndolas sin que el intríngulis haya desaparecido por eso.
De seguro las técnicas histológicas de 2006 se benefician con recursos
físicoquímicos inexistentes hace noventa años, pero no suplen a la ardua labor
y, con ella, las sencillas técnicas clásicas tienen todavía mucho que
proporcionar a los investigadores provistos del adecuado panorama
neurobiológico, histopatológico y filogenético. Un buen ejemplo fue la
presentación que en 1998 hizo Francis Crick a la Academia Nacional de Ciencias de los Estados Unidos, transmitiendo el
descubrimiento de varios investigadores de "A neuronal morphologic type unique to humans and great apes"
logrado trabajando solamente con el método de Nissl (esto fue luego publicado
en los Proceedings Vol. 96 Nro. 9
pág. 5268-5273, el 27-IV-1999).
En el 50° aniversario de la muerte de Christofredo
Jakob he querido rendir este sencillo homenaje tratando otra vez uno de los temas
que más lo apasionó en su dilatada existencia científica: el origen
ontofilogenético de la corteza cerebral, que lo llevó a fundar su teoría de la
doble corteza (Doppelrinde). En este
estudio se ven las dificultades del problema, los hombres que lo acompañaron o
precedieron en sus ideas y la importancia que, ya en este año 2006, el tema mantiene
todavía.
Bibliografia
1. BEVAN LEWIS, W. y CLARKE, H.: On the cortical lamination of
the motor area of the brain. Proc. Roy.
Soc. 27, 38, 1878.
2. JAKOB, CHR.: La célula cortical en la locura. Anales de la Administrac. Sanatorial y
Asist. Pública, 1910, pág. 5.
3. JAKOB, CHR.: Vom Tierhirn zum Menschenhirn I. Teil. Tafelwerk nebst
Einführung in die Geschichte der Hirnrinde. Lehmann, München, 1911.
4. JAKOB, CHR.: Das Menschenhirn (eine Studie über den Aufbau and die Bedeutung
seiner Grauen and Rinde) I. Teil: Tafelwerk nebst Einführung in den
Organisationsplan der menschlichen Zentralnervensystems. Lehmanns, München,
1911.
5. JAKOB, CHR.: Ueber die Ubiquität des sensomotorischen Doppelfunktion der Hirnrinde als Grundlage
einer neuen biologischen Auffasung der kortikalen Seelenorgans. Münchener Medizinischen Wochenschrift 9,
1912.
6. JAKOB, CHR. y ONELLI
CL.: Atlas del cerebro de los
mamíferos de la República
Argentina (Estudios
anatómicos, histológicos y biológicos
comparados sobre la evolución
de los hemisferios y de corteza cerebral) Kraft, Bs. As., 1913.
7. JAKOB, CHR.: La
filogenia cortical. Sobre la corteza
cerebral de gimnofiones y
amfisbenas argentinas. Actas y trabajos del 1er. Congreso
Nacional de Medicina,
Bs. As., 1918, 4, 82.
8. KAPPERS, C. Ü. ARIENS: The phylogenesis of the paleocortex
and archicortex compared with the evolution of the visual neocortex. Arch. of Neur.
and Psychiat, 1909, 1, 161.
9. KUHLENBECK, H.: Über den Ursprung der Grosshirnrinde. Anat. Anz., 55, 337-365, 1922.
10. LUYS, J.: "Le Cerveau", E. Martinet, Paris, 1876.
11. MEYNERT, TH.: Der Bau der Gross-Hirnrinde und seine örtlichen
Verschiedenheiten nebst einem pathologisch-anatomischen Corollarium. Vierteljahresschrift. f. Psychiat. 1,
77-93 y 198-217 (1867) y 88-113 (1868).
12. MEYNERT, TH.: Neue Untersuch. ü. den Bau der Grosshirnrinde. Osterr. Med. Jahrb. 17, 1869.
13. MOTT, F. W.; The progressive Evolution of the structure
and functions of the visual cortex in mammalia. Archives of Neurology, 3. 1907.
14. MOYANO, B.: Christfried Jakob. Acta
Neuropsiquiat. Arg. 1957, 3,
109.
15. NISSL, F.: Die Neuronenlehre und ihre Anhänger, G. Fischer, Jena, 1903.
16. NISSL, F.: Experimental Ergebnisse zur Frage der Hirnrindeschichtung, Mschr. Psychiat. 23, 1907.
17. NISSL, F.: Zur Lehre u. Lokalisation der Grosshirnrinde des Kaninchens Akad. Wiss Math. Naturw. Kl. (Sitzgsber. Heidelberg) Bd. II, 3, 1911.
18. RAMON y CAJAL, S.: Estudios
sobre la corteza cerebral humana. R. Trim. Micrográf. IV, 1899. Bressler tradujo estos estudios al alemán y así publicó en Leipzig los
volúmenes de la Trimestral Micrográfica
desde 1899 hasta 1901 (Amb. Barth, 1906), con el título Studien über die Hirnrinde
des Menschen.
19. SPATZ, H.: Nissl und die theoretische Hirnanatomie. Arch. f. Psychiat. 87, 1929.
Copyright © 2006 Electroneurobiología. Esta investigación original constituye un artículo de acceso público; su
copia exacta y redistribución por cualquier medio están permitidas bajo la condición
de conservar esta noticia y la referencia completa a su publicación incluyendo
la URL original (ver arriba). / This is an Open Access article: verbatim
copying and redistribution of this article are permitted in all media for any
purpose, provided this notice is preserved along with the article's full
citation and original URL (above).

revista
Electroneurobiología
ISSN: 0328-0446
Croquis de este sitio - Outline of this site
Índice - Table des matières
- Inhaltsverzeichnis - Table of Contents
Some
downloadable articles and documents in this site are in PDF format (.pdf). To
view or print these articles, you must have Adobe Acrobat Reader installed. Acrobat Reader can be downloaded from
Adobe's web site free of charge by clicking here / Se recomienda mucho
leer o imprimir algunos de los trabajos mas extensos con el programa Acrobat
Reader,
que puede obtenerse gratuitamente pulsando aqui ![]()
2006 – CINCUENTENARIO DE LA MUERTE DE CHRISTOFREDO JAKOB – 2006
2006 – CENTENARIO DEL NACIMIENTO DE BRAULIO MOYANO – 2006
2006 – Año de homenaje al Dr. Ramón Carrillo – 2006
en el quincuagésimo aniversario
de su deceso
Decreto 1558/2005 de la Presidencia de la Nación
Ver debajo las publicaciones
concernientes al mismo
2006 – A TREINTA AÑOS DE LA PATENTE BRITÁNICA 1.582.301 – 2006

Haga doble
"click" en el título de cualquier artículo, para leerlo ahora -
Double-click on any article to read it now:
SOCIOLOGÍA DE LAS NEUROCIENCIAS
![]() Pour comprendre
l'enjeu : L’anthropologie ganglionnaire, un psychovirus démasqué (français)
Pour comprendre
l'enjeu : L’anthropologie ganglionnaire, un psychovirus démasqué (français)
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (190 kB:
recomendada) o .DOC (76 kB).
![]() L'aliénisme en
Argentine : Diego Alcorta (1827) : Dissertation sur la manie... aiguë?
(français)
L'aliénisme en
Argentine : Diego Alcorta (1827) : Dissertation sur la manie... aiguë?
(français)
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (600 kB: recomendada)
o .DOC (320 kB).
![]() Metaphors at odds in conceiving organismal-societal government: The Political Structure of the Brain: Cerebral Localization in Bismarckian Germany
(English)
Metaphors at odds in conceiving organismal-societal government: The Political Structure of the Brain: Cerebral Localization in Bismarckian Germany
(English)
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (228 kB: recomendada)
o .DOC (175 kB).
ELECTRONEUROBIOLOGÍA
Efectos
relativísticos en biofísica cerebral:
Puede leer, imprimir o guardar en su disco duro esta investigación en versión .PDF (496 kB: recomendada) o .DOC (227 kB)
SUMARIO Y
PÁRRAFOS INICIALES EN CASTELLANO
Diversificación
de recursos electroneurobiológicos en la evolución del sistema nervioso:
Puede leer, imprimir o guardar en su disco duro esta investigación en versión .PDF (733 kB: recomendada) o .DOC (406 kB)
auch als .PDF (285 kB) oder .DOC (161 kB)
Puede leer, imprimir o guardar en su disco duro esta investigación
en versión .PDF (354 kB:
recomendada) o .DOC (236 kB).
Cálculo
de potenciales dentro de las células
 Calcule
intensidades eléctricas y magnéticas en cada compartimiento neuronal: The nervous principle: active versus passive electric
processes in neurons (Explains how to calculate electric and magnetic
field strengths inside different neuronal compartments) (LONG FILE IN ENGLISH with
Bulgarian, Russian and Spanish abstracts/TOCs)
Calcule
intensidades eléctricas y magnéticas en cada compartimiento neuronal: The nervous principle: active versus passive electric
processes in neurons (Explains how to calculate electric and magnetic
field strengths inside different neuronal compartments) (LONG FILE IN ENGLISH with
Bulgarian, Russian and Spanish abstracts/TOCs)
Podrá leer, imprimir o guardar en su disco duro esta investigación en versión .PDF (2 Mb): recomendada) o .DOC (1,5 Mb). También como .html comprimido (compressed .HTML folder: 0,5 Mb) .ZIP.
Evaluación de
potenciales fuera de las células
Signal analysis to exploit the information of steady-state recordings: Do’s and don’ts in Fourier analysis of steady-state potentials
(Assumptions in the discrete Fourier transform (DFT) not necessarily fulfilled in real-world applications) (English)
NOCIONES GENERALES
Conceptos:
![]() Noticia general -- ¿Qué es electroneurobiología? -- La
atmósfera intelectual (all in Spanish) -- Main Technical Ideas / Conceptos
técnicos principales (English and
Spanish) -- El descubrimiento de la Doppelrinde (German and Spanish)
Noticia general -- ¿Qué es electroneurobiología? -- La
atmósfera intelectual (all in Spanish) -- Main Technical Ideas / Conceptos
técnicos principales (English and
Spanish) -- El descubrimiento de la Doppelrinde (German and Spanish)
![]() Comentando una "ilusión óptica" / Commenting an "optical illusion":
A visual yet non-optical subjective intonation:
una entonación subjetiva visual pero no óptica
(English and
Spanish)
Comentando una "ilusión óptica" / Commenting an "optical illusion":
A visual yet non-optical subjective intonation:
una entonación subjetiva visual pero no óptica
(English and
Spanish)

![]() UNA EXPLICACIÓN ESENCIAL:
UNA EXPLICACIÓN ESENCIAL:
Puede obtener un
archivo .PDF (401 kB:
recommended) o .DOC (217 kB)
para imprimir este trabajo.
Historia de las
experimentaciones:
Table of Contents (partial) of "Sensing: a new
fundamental action of nature" (English) -- Índices
Recepción
de los aportes de Chr. Jakob en la neurobiología germana
You can download a .PDF (700 kB:
recommended) or .DOC (1.3 MB)
file for printing this work
You can download a .PDF (673 kB:
recommended) or .DOC (1.29 MB) file
for printing this work.
![]() Adolf von Strümpell (1853-1926) fue factor definitorio en la ruptura de Josef Breuer con Sigmund Freud. Los vínculos de amistad que unían a Christofredo Jakob (1866-1956) con su ilustre maestro y buen amigo nunca se resintieron por la separación que les impusiera la distancia y el tiempo; Jakob, asimismo, respetó el secreto de von Strümpell acerca de la enfermedad que padecía Lenin. -- Oddo y ot.: El Maestro de la medicina platense Christofredo Jakob, discípulo y amigo de Adolf von Strümpell
(Spanish: ARCHIVO DE DESCARGA LENTA POR LLEVAR MUCHAS ILUSTRACIONES)
Adolf von Strümpell (1853-1926) fue factor definitorio en la ruptura de Josef Breuer con Sigmund Freud. Los vínculos de amistad que unían a Christofredo Jakob (1866-1956) con su ilustre maestro y buen amigo nunca se resintieron por la separación que les impusiera la distancia y el tiempo; Jakob, asimismo, respetó el secreto de von Strümpell acerca de la enfermedad que padecía Lenin. -- Oddo y ot.: El Maestro de la medicina platense Christofredo Jakob, discípulo y amigo de Adolf von Strümpell
(Spanish: ARCHIVO DE DESCARGA LENTA POR LLEVAR MUCHAS ILUSTRACIONES)
Puede obtener un
archivo .PDF (2,5 MegaBytes:
VERSIÓN RECOMENDADA) o .DOC (6 MegaBytes)
para imprimir este trabajo.
Puede obtener un
archivo .PDF (256 kB:
recommended) o .DOC (106 kB)
para imprimir este trabajo.
Recepción
de los aportes de Chr. Jakob en la neurobiología y la sociedad argentina
Puede obtener un archivo .PDF (277 kB:
recommended) o .DOC (170 kB)
para imprimir este trabajo.
![]() Piva y Virasoro - Christofredo
Jakob, neurobiólogo: científico en diálogo filosófico (Spanish)
Piva y Virasoro - Christofredo
Jakob, neurobiólogo: científico en diálogo filosófico (Spanish)
Puede obtener un archivo .PDF (391 kB:
recommended) o .DOC (466 kB)
para imprimir este trabajo.
Puede obtener un archivo .PDF (385 kB:
recommended) o .DOC (813 kB)
para imprimir este trabajo.
Puede obtener un archivo .PDF (384 kB:
recommended) o .DOC (814 kB)
para imprimir este trabajo.
Panorama
evolutivo:
FILOSOFÍA DE LA CIENCIA - CONCEPTO DE TIEMPO EN
NEUROBIOFÍSICA

Why is
Time Frame-dependent in Relativity? Minkowski's spacetime as a Kantian
'condition of possibility' for relativistic calculations (English) -- Abstract: Minkowski spacetime is a condition
of possibility for relativistic calculations. It keeps special relativity
kinematic (i.e. avoids introducing it it causal dynamics) but in exchange
forces relativistic observers to describe at once past, present, and future
states along the length of the observed moving entities. This has been
considered a proof that past and future components of real entities enjoy an
unremitting mode of existence, because, if the world described by relativity
were three-dimensional, the kinematic consequences of special relativity and
the experiments confirming them would be impossible. The latter is acknowledged
as exact but the assumed proof is viewed as incorrect because, such imposition
being a Kantian condition of possibility, it cannot reveal anything about what
Kant called noumenon, namely extramentality. ( Traducción castellana en preparación ).
You can download a .PDF (1 MB: recommended) or .DOC (571 kB) file for printing this work.
RESUMEN
DIVULGATORIO Y PARA ESTUDIANTES EN CASTELLANO
Puede leer, imprimir o guardar en su disco duro el resumen en
castellano en versión .PDF (204 kB:
recomendada) o .DOC (96 kB)
“ANTAGONISMO ENTRE
CIENCIAS DURAS Y HUMANIDADES BLANDAS”
MALFORMACIONES Y PAPEL DEL ÓRGANO CEREBRAL
![]() Christofredo Jakob: “Los
Monstruos Anencéfalos” (Spanish)
Christofredo Jakob: “Los
Monstruos Anencéfalos” (Spanish)
Puede leer, imprimir o guardar en su disco duro esta
investigación en versión .PDF (346 kB:
recomendada) o .DOC (280 kB).
BIOÉTICA
![]() Éthique de la Bio-Éthique (français) ¡Nuevo!
Éthique de la Bio-Éthique (français) ¡Nuevo!
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (323 kB: recomendada) o .DOC (161 kB).
EL PRESUNTO
DUALISMO CUERPO - ALMA
Puede leer, imprimir o guardar en su disco duro esta investigación en
versión .PDF (373 kB:
recomendada) o .DOC (201 kB)
PSICOANÁLISIS Y
FACILITACIóN PSICOSOMÁTICA DE
LA ENF. DE ALZHEIMER
Puede leer, imprimir o guardar en su disco duro esta investigación en
versión .PDF (502 kB:
recomendada) o .DOC (110 kB)
PERICIAS
JUDICIALES Y CASUÍSTICA
Puede leer, imprimir o guardar en su disco duro esta investigación en
versión .PDF (300 kB:
recomendada) o .DOC (150 kB)
NUESTRA GENTE
![]() Arturo Carrillo, con la colaboración de Augusto Raúl Carrillo : segmentos de su libro "Ramón Carrillo. El hombre... El médico... El sanitarista"
(Spanish)
Arturo Carrillo, con la colaboración de Augusto Raúl Carrillo : segmentos de su libro "Ramón Carrillo. El hombre... El médico... El sanitarista"
(Spanish)
Libro declarado de Interés Cultural por la Legislatura de la Ciudad Autónoma de Buenos Aires
![]() seguidos de una
Noticia biográfica del Dr. Arturo Carrillo (Spanish)
seguidos de una
Noticia biográfica del Dr. Arturo Carrillo (Spanish)
Puede obtener un archivo .PDF (2,45 MB:
recommended) o .DOC (2,1 MB)
para imprimir estos dos trabajos.
También puede descargar esos archivos comprimidos, como.PDF.ZIP (2,1 MB:
recommended) o .DOC.ZIP (1,7 MB)
para guardar o imprimir posteriormente estos artículos.
_____________________________
![]() Breve reseña biográfica: Ramón Carrillo, el Gran
Sanitarista Argentino, por Marcos Ordóñez (Spanish)
Breve reseña biográfica: Ramón Carrillo, el Gran
Sanitarista Argentino, por Marcos Ordóñez (Spanish)
ESCATOLOGÍA – POSTMORTALIDAD – EL PUESTO DE LA HUMANIDAD EN LO REAL
(ESCHATOLOGIE – LEBEN NACH DEM TOD – DIE STELLUNG
DES MENSCHEN IM KOSMOS)
A Palindrome: Conscious Living Creatures as Instruments of Nature; Nature as an Instrument of Conscious Living Creatures. Cuando tanto materialistas como idealistas nos describen todas las cosas tomadas en conjunto, hoy los científicos les podemos replicar: "Sé verlas al revés" (English).
[Nota para distraídos: "Sé verlas al revés" es también un palindrome]
SUMMARY: It is reported a palindromic relationship between the astrophysical-biological evolution and the experiencing beings in it. The issue is related with ascertaining if nature is an instrument (as merely a means), instead of having any intrinsic value (an end in itself); and, likewise, if conscious beings are merely a means (one to entropize nature faster) or either possess any intrinsic value. Two possibilities are deemed not indifferent in this regard: either reading the whole set of empirically-found realities or facts makes sense in both directions (palindromic reading of nature), or, rather, that sense can only be ascribed to such a set by reading it in some single direction. A single direction means reading nature in a classic, materialist or idealist sense; both directions' sense means a mirror or reciprocal functionalization, in which each of both realities (mind-possessing living creatures, and astrophysical-biospheric evolution) uses for its own ends the reality that uses it as a means. At stake, therefore, is establishing if axiological readings ascribing a sense to what is found going on in the universe can be obtained in both directions, or not. On this alternative, it is claimed, pivots the possibility of ascertaining, e.g., whether conscious beings are worthier than non-conscious nature, or not - a topic assumed consequential for philosophy, ecology, ecofeminism and biocentric environmental movements, and ethics.
You can download a .PDF (675 kB: recommended) or .DOC (310 kB) file for printing this work.
PROCEDIMIENTO
![]() Los "Cortes de Jakob": Christofredo Jakob: La técnica moderna en la autopsia del cerebro. Exposición clara y didáctica de la técnica, por su autor, con ilustraciones y la explicación de sus fundamentos (Spanish).
Los "Cortes de Jakob": Christofredo Jakob: La técnica moderna en la autopsia del cerebro. Exposición clara y didáctica de la técnica, por su autor, con ilustraciones y la explicación de sus fundamentos (Spanish).
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (2 MB:
recomendada) o .DOC (1,6 MB).
Estos artículos cubren sólo algunos de los temas que se explorarán en este documentado sitio de Red, por ahora en construcción. Abrirá completo en unos meses.

These texts cover just some of the topics that will be explored at this extensive website which is now under construction. In a few months it will open in full.
Introducción (English
opening below): Mientras las
neurociencias, como rama de la investigación biomédica, son una herramienta
para combatir la enfermedad, como rama de las humanidades son una herramienta
para sondear la condición humana. Por ser ambas cosas, las neurociencias
aportan una descripción de la gente cuya veracidad se tiene por fundada en la
eficacia de esas neurociencias para sugerir remedios contra las enfermedades.
Esta eficacia hace creíble su descripción. Tal como cualquier autorretrato
creíble, asimismo la autodescripción neurocientífica induce a la gente a
aceptar o rechazar ciertas perspectivas sociales y medidas de gobierno – por
donde la descripción neurocientífica de la gente también se torna herramienta
política. Ello ocurre de modo similar a como algunos intereses se benefician y
otros se perjudican con las ideas acerca de la condición humana propagadas por,
digamos, el subjetivismo de The Matrix o la negación gnosticista por “Harry
Potter” de ciertos límites en nuestras capacidades. La electroneurobiología es
la parte de las neurociencias que estudia dentro del tejido cerebral las
fluctuaciones, orquestadas biológica o psicológicamente, de cierto campo
físico, a saber el campo eléctrico, que ya es bastante bien conocido por otras
áreas de la tecnociencia contemporánea. Este campo eléctrico se superpone con
otro campo físico y las fluctuaciones de ambos se acoplan intimamente; el
segundo es aquél campo físico en cuyas diferentes parcelas la gente encuentra
más precisamente localizados en la naturaleza sus intercambios causales. Por
cuanto aún no podemos manipular ni sondear de otra manera este último campo –
tal como también ocurre, por ejemplo, con el campo gravitatorio – nuestro único
acceso a este segundo campo es el acople electroneurobiológico con el mismo.
Debido a esta circunstancia la electroneurobiología se torna cardinal para los
roles científico, humanístico y político para los que se instrumentan las
neurociencias. La revista Electroneurobiología, fundada en 1993 como
suplemento especializado de la Folia Neurobiológica Argentina iniciada
en 1939 por C. Jakob, cubre temáticamente todos estos roles de su disciplina:
tanto las cuestiones humanísticas cuanto las atingentes a las ciencias
biomédicas y a la pragmática sociopolítica.
Este sitio de red presenta versiones completas y de
acceso público de una selección de los artículos de Electroneurobiologia.
Estos pues tratan los temas o áreas aludidos por las palabras clave listadas al
pie de esta Introducción. Algunas de esas palabras están en diversos idiomas,
tal como se las emplea en ciertos artículos. Deseamos y esperamos ser capaces
de presentar los trabajos de investigación con al menos los resúmenes en los
idiomas oficiales de los países del Mercosur – castellano, portugués y guaraní
–así como en otros lenguajes tradicionales de la ciencia como francés, alemán e
inglés y algunos otros de culturas no occidentales. Pero mantener en línea Electroneurobiologia
no es ni el único ni el principal esfuerzo en que estamos comprometidos los
científicos de este Laboratorio. Será bienvenida toda ayuda para traducir hacia
cualquier idioma los resúmenes o, como ideal, los artículos. Incluso esbozos
preliminares de esas traducciones serían muy útiles, ya que nosotros luego
podríamos procurar refinarlos. Si usted aprecia la importancia de estos
resultados investigativos y puede y quiere ayudar a comunicarlos con traducciones
o esbozos de ellas hacia cualquier idioma, por favor contáctenos en el correo
electrónico indicado arriba.
Nota sobre neuromercadeo ("neuromarketing") y neuroeconomía (agregada en diciembre de 2005): Cierto número de lectores llegan ahora a esta revista en búsqueda de ideas técnicas útiles para juzgar acerca del neuromercadeo y la neuroeconomia. Mientras que no nos es posible dedicar los esfuerzos adecuados para esclarecerles las insostenibles simplificaciones (a veces llamadas "neuromacaneo", neurobabble) rampantes en la mayoría de los tratamientos de dichos temas, puede serles útil apuntar aquí una breve evaluación, a guisa de conclusión. Varios artículos de esta revista sirven a clarificar por qué las imágenes de resonancia magnética nuclear funcional, o fMRI, que miden cambios del flujo sanguíneo cerebral para determinar la intensidad promedio de la actividad que genera ondas electroencefalográficas en diferentes áreas del cerebro, no pueden penetrar en el psiquismo.
La fMRI, antes bien, lleva a cabo solamente lo mismo que realiza la observación con moderno instrumental de cualquier otra conducta; esto es, describe con precisión algún aspecto de dicha conducta corporal, de modo de permitir al observador conjeturar mejor, siempre en base a su experiencia, la motivación psicológica de tal conducta – si es que cabe suponerle alguna. Las ondas eléctricas del tejido cerebral, lejos de constituir "la materia del pensamiento humano", son los "movimientos" del estado eléctrico cerebral – similares a los movimientos de un miembro, que también pueden ser voluntarios o involuntarios – por los cuales el órgano cerebral reacciona bien sea a ocurrencias extramentales o bien a iniciativas mentales.
Los psiquismos siguen siendo privados: persuadibles, quizás, pero incompartibles e inauguradores de cadenas causales originadas por decisiones privadas. En la precisa medida en que la investigación del consumo ("consumer research") impropiamente describe a los psiquismos como si fueran puramente reaccionantes, a las memorias como si fueran marcas en el cerebro, y a la atención como si solamente fuera movida por su objeto atendido, la maniobra de enseñar por todo el globo a las empresas cómo comunicarse eficaz e instantáneamente con el "botón de compra" ("buy button"), esa "parte del cerebro donde la decisión de adquirir tiene lugar", y así poner a los compradores en un estado de "dopamina alta" que desactive el "hemisferio analítico del cerebro", es sólo una manera de que tales enseñantes se construyan laboratorios de fachada para seguir sacándole dinero a las empresas – no de que estas últimas aumenten sus ingresos.
![]()
English opening: While neuroscience, as a branch of biomedical research, is a tool for
coping with disease, as a branch of humanities it is a tool for fathoming the human
condition. And because of its being both, neuroscience conveys a description of
people whose truth is held grounded upon the neuroscience’s effectiveness to
suggest remedies for disease. This effectiveness makes it believable. Like any
believable selfdepiction, the neuroscientific selfportrait induces people to
agree or disagree with specific societal views and governmental measures –
whereby the neuroscientific description of people also becomes a tool for
politics. It happens in a way similar as some interests benefit and others are
countered by ideas about the human condition vehicled through, say, The
Matrix’s subjectivism or “Harry Potter”’s gnosticist denial of certain
limits to one’s powers. Electroneurobiology is the part of neuroscience
studying within the brain tissue the biologically or psychologically
orchestrated fluctuations of certain physical field, namely the electric field,
that stands quite well known from other areas of contemporary technoscience.
The electric field overlaps and is intimately coupled with another physical
field, into different parcels of which field people find their causal
interactions the most immediately localized in nature. Since we cannot yet handle
or otherwise probe this later field – as it too happens, for example, with
regard to the gravitatory one – our sole current access to this later field is
the electroneurobiological coupling with it. Thereby electroneurobiology
becomes pivotal for the scientific, humanistic and political roles accomplished
by neuroscience. The journal Electroneurobiologia, founded in 1993 as a
specialized supplement of the 1939-started Folia Neurobiólogica Argentina,
covers all these roles of its branch of learning: the humanistic roles as well
as the ones in biomedical science and political affairs.
This website presents open-access, full-text versions of selected
articles of Electroneurobiologia,
thus dealing with the themes or areas referred to by the keywords recorded
at the end of this notice – some are non-English words, as they are used in
some articles. We wish and hope to be able of presenting the research papers
with at least the abstracts in the official languages of the Mercosur countries
– Spanish, Portuguese and Guarani – as
well as other traditional languages of Western science such as French, German
and English and some others of non-Western cultures. Yet keeping Electroneurobiologia
online is not the sole or the main effort in which the Laboratory’s scientists
are engaged. Any help to translate the abstracts or, ideally, the articles into
any language is welcome. Even preliminary drafts of such traslations are very
useful, as we can then procure to refine them. If you appreciate the importance
of these research results and can and wish help with translations or drafts
into whatever language, please contact us at the above e-mail.
Note on neuromarketing and neuroeconomics (added in December, 2005): A number of people nowadays come to this journal looking for technical insights useful to judge neuromarketing and neuroeconomics. While we cannot devote efforts to clarify for such special-purpose readers the untenable simplifications (not seldom called "neurobabble", neuromacaneo) rampant in most treatments of those fields, a brief conclusive assessment may be of help and is in order here. Several articles of this journal show why functional magnetic resonance imaging, or fMRI, which measures changes in blood flow in order to ascertain the average intensity of brain-wave generating activity in different areas of the brain, cannot delve into the mind.
fMRI rather does just the same as the instrumental observation of any other behavior, i.e. accurately depicts some aspects of this bodily behavior so as to allow the observer to guess its psychological motivation – if any at all. Brain waves, far from being "the stuff of human thought", are the "movements" of the brain's electrical state – like as the movements of a limb, which also may be voluntary or involuntary – whereby the brain organ reacts either to extramental events or to mental initiatives.
Minds remain private: persuasible, maybe, but unshareable and inaugurating causal chains on private decisions. Inasmuch as consumer research improperly describes minds as purely reactive, memories as cerebral marks, and attention as driven solely by the object, the endeavor, of teaching companies around the world how to communicate strongly and instantly to the "buy button", that "part of the brain where the purchase decision takes place", and causing the 'analytical side to fall away' by putting shoppers into a "high dopamine" state, is a way for the teachers to build facade laboratories and go on making money from the companies – not for the latter to increase revenues.
[Prof. Mariela Szirko
cuida estas páginas/cares of these pages.]
![]()