Gobierno de la ciudad de Buenos Aires
Hospital Neuropsiquiátrico
"Dr. José Tiburcio Borda"
Laboratorio de Investigaciones Electroneurobiológicas
y
Revista
Electroneurobiología
ISSN: ONLINE 1850-1826 - PRINT 0328-0446

Vías nerviosas:
neurones y neurofibrillas
por
Matilde
V. Villa
Profesora Normal
Contacto
/ correspondence: vixit
con notas editoriales de
Mariela
Szirko
Electroneurobiología Vol. 4 (3), agosto 1997, pp. 1-56; URL <http://electroneubio.secyt.gov.ar/index2.htm>
Originariamente publicado en la revista El Libro, Vol. 1, pp. 524-547 y 638-649,
1907 y Vol. 2, pp. 75-85, 1908.
Copyright
© 1997 Electroneurobiología. Este
trabajo es un artículo de acceso público; su copia exacta y redistribución por
cualquier medio están permitidas bajo la condición de conservar esta noticia y
la referencia completa a su publicación incluyendo la URL (ver arriba). / This
is an Open Access article: verbatim copying and redistribution of this article
are permitted in all media for any purpose, provided this notice is preserved
along with the article's full citation and URL (above).
Imprimir este archivo no
preserva el formato ni la paginación original. Puede obtener un archivo .PDF
(recomendado: 917 kB) o .DOC
(556 kB) para leer o imprimir
este artículo, desde aquí o de / You can download a .PDF
(recommended: 917 kB) or .DOC
(556 kB) file for reading or printing, either from here or <http://electroneubio.secyt.gov.ar/index2.html
_______
SUMARIO:
-¿Qué se
entiende por neurón?
-Comunicación
de los neurones
-Concepciones
antiguas
-Teoría
de Gerlach
-Teoría
de Golgi
-Concepciones
nuevas
-Críticas
acerca de estas teorías
-Objeciones
antiguas
-Objeciones
nuevas.
Noticia preliminar, por Mariela Szirko. Hace noventa años la revista del profesorado
argentino (vol. 1, pp. 524) brindaba esta noticia: "En el
siguiente número de “El Libro” se publicará la segunda parte de éste importante
trabajo en que se resume todo cuanto se ha escrito sobre los neurones
y neurofibrillas, temas de palpitante actualidad." El valor de esta obra de una de las alumnas de Christofredo
Jakob radica en que revela y marca las aspiraciones docentes argentinas. Se
inserta en la misma línea que la obra apenas posterior de Jakob (1915-1918), Tratado
de Biología General y Especial para el uso de la enseñanza elemental, secundaria
y superior en la República Argentina (vols. I –Kraft, Buenos Aires, 1915,
pp. 1-650– y II, Fascículos I-IV, 1917-1918, pp. 651-854: Kraft, Buenos Aires,
y Revista del Jardín Zoológico, separatae), de la que hace muy poco se han
destruído con saña unos seis mil ejemplares conservados como nuevos y aún
útiles. Útil contraste este; revés que estimula, gotícula de una catarata que
cuanto más aplasta más fuerte remueve y alza al topar con el fondo de piedra.
Por si alguna imprecisión cupiera acerca de las
aspiraciones que mueven la pluma de Matilde Villa y su entorno, y para
renovarlas por si alguna vez perdiéramos de vista la roca y nos sonrojara vocearlas,
transcribo, de otra nota publicada por la misma revista cerca de su trabajo, algunas
líneas al azar: "Y lo que tú serás, Argentina, yo lo vislumbro.
Para ti el porvenir amplio.
Risueño, como es amplia tu dilatada pampa, como risueñas son tus provincias
ribereñas; para ti
la
vida exuberante fastuosa, como exuberante es tu naturaleza y fastuosa la
majestad andina. Para ti el mañana alegre, placentero; ya que en tu
regazo habrán cristalizado todos los ensueños, todas las esperanzas, los amores
todos de todas las razas que aquí llegan ansiosas de cooperar a tu crecimiento
y desarrollo. Bendita, bendita tú, Argentina, que vas dando vida real y poco a
poco a un colosal ensueño, el de la fraternidad universal, ya que bajo tu
bandera blanca como la alegría y azul como la risueña bóveda celeste, se
estrechan amorosamente las manos, así los nacidos en la poética Italia como
los que vieran la luz en la nebulosa Albión, así los hijos de la republicana
Francia como los del imperio germánico: así los que aún ven envuelto su
cerebro con las brumas de la ignorancia como los que sienten agitarse en su
mente los hervores de fecundante idea. Para ti, el amor
idólatra de tus hijos; para ti, Argentina, los nobilísimos amores de los extranjeros
en tu suelo radicados."
Por esos anhelos que corporiza, este trabajo – que preserva
aquí su ortografía original, para recordarnos de qué época procede – nunca
perderá su valor histórico. Continuará siempre exhibiendo los ideales de una
patria que, entre otros elementos, ha de hacerse vigorosa por sus propósitos, como
aquel ya manifiesto en 1907 de llevar la explicación de temas científicos de
palpitante actualidad a los estudiantes del ciclo secundario, la mayoría de
entre doce y diecisiete años de edad. Pero si no se le piden peras al olmo, en
este caso si no se espera que la presente sinopsis nos hable de neuroquímica,
expresión de genes o microbiofísica, este artículo no se conforma incluso con
eso. Es tal la pertinencia de las observaciones comentadas y la profundidad
conceptual de su síntesis que aún ofrece utilidad docente y abundante materia
para la reflexión técnica, obligando por ejemplo a confrontar si el hecho de
tener que describir al tejido nervioso como carente de sustancia intercelular
tiene por causa su función, o su historia evolutiva, o nuestra idea de la
sustancia intercelular como algo tangible que excluye las conformaciones
electroneurobiológicas de campo. (1)
________
(1)
Cf.
Ávila, Alicia, y Crocco, Mario (1996), Sensing: A New Fundamental Action of
Nature (Folia Neurobiológica Argentina, vol. X: Institute for Advanced
Study, Buenos Aires), Chapter 1.5, pp. 432-490: The Three Encaptical Functions
Physically Accomplished by Sense-interacting Parenchymæ (Secciones: §1.5.1:
Scale anomaly of the sensed foam’s galvanic skeleton, p. 432; §1.5.2: Function
1 — Shaping the structures of the neurocognitive contents; p. 435; Conative
imagining in mnesic evocation, p. 438; §1.5.2.1: Field skeleton of the
introspected condensed matter, p. 449; §1.5.2.2: Contents and operations of the
field skeleton, p. 451). Véase
también Crocco, Mario y Contreras, Norberto (1986), "El esqueleto de
campos electromagnéticos del tejido neurocognitivo: un nuevo concepto del
órgano cerebral", La Semana Médica 168 (4), enero 26; Crocco, Mario
(1984), Cómo el tejido neurocognitivo
genera fenómenos psicológicos (Cát. Anatomía y Fisiol. Sistema Nervioso,
Carrera de Psicología, Univ. de Buenos Aires).
NEURONES Y NEUROFIBRILLAS — I
La
ciencia, como dice Burdach, no es una simple acumulación de hechos aislados e
invariables, sino por el contrario es un conjunto
de hechos, que día á día va aumentando de extensión por los nuevos hechos que
van surgiendo y que es necesario agregarlos a los que nos son ya conocidos.
Estos
hechos nuevos vienen a menudo a llenar las lagunas que existen en el terreno
científico ó bien a variar el curso de nuestras ideas y, por lo tanto, la
dirección de nuestros estudios.
Tal
cosa acontece en todas las ciencias; pero su influencia es aún más determinante
en las ciencias nuevas que están en vías
de formación, verbigracia: la Psicología.
En
efecto, esta ciencia de vastísimos horizontes es aún nueva para nosotros, y sus
variados estudios son todavía objeto de delicadas y admirables investigaciones.
Entre
los estudios que abarca dicha ciencia uno de los más interesantes es el estudio
del sistema nervioso.
Veamos
algunas consideraciones acerca de este estudio:
El
hombre, considerado anatómicamente, no es más que el conjunto de un sinnúmero
de células de naturaleza distinta según sean las funciones que están llamadas
á desempeñar.
Estas
células sufren las transformaciones necesarias según las funciones que
desempeñan; y luego se unen las que pertenecen á una misma categoría, formando
así aglomeraciones en la sustancia intercelular, ó bien, por fusión,
constituyen masas ó complejos más ó menos considerables, denominados tejidos.
Estos
tejidos varían entre sí, ya sea por la naturaleza de las células que los forman
ó ya por la sustancia intercelular; de ahí la necesidad de agruparlos en distintas
categorías: á la primera categoría pertenecen los tejidos formados por células
simples y por sustancia intercelular liquida y abundante; á la segunda categoría
pertenecen los tejidos cuyas células han sufrido una transformación parcial y
cuya sustancia intercelular es más ó menos sólida, y á la tercera categoría
pertenecen los tejidos cuyas células han sufrido ya una transformación total
y que carecen de sustancia intercelular.
A
esta última categoría pertenece el tejido muscular y el tejido nervioso.
Este
último tejido es el encargado de recibir las impresiones para transformarlas
luego en sensibilidad ó movimiento y viene a ser, por lo tanto, el agente
principal de nuestros actos voluntarios. Consta de dos elementos esenciales:
las fibras y las células nerviosas.
La
célula nerviosa con sus diferentes prolongaciones desde el doble punto de
vista. anatómico y funcional forma un todo continuo, un todo indivisible, una
verdadera unidad nerviosa á la cual Waldeyer denomina neurón.
Este
término de neurón es considerado hoy como término clásico y, como nos hemos de
ocupar continuamente de él, consideramos necesario definir primero qué es lo
que se entiende por neurón: «El neurón no es más que una célula nerviosa con
todas las prolongaciones que emanan de ellas».
Luego,
si no es más que una célula nerviosa, distinguiremos en todo neurón tres partes
esenciales: primero una parte central (que no es más que la célula propiamente
dicha) que se denomina cuerpo celular, el cual comprende el núcleo con sus
correspondientes nucleolos; segundo: una parte periférica constituida por las
prolongaciones protoplasmáticas y sus diversas ramificaciones; tercera: una
segunda parte periférica constituida por las prolongaciones cilindroejes
cualquiera que sea su extensión, como la función que desempeñe, pudiendo
hallarse en un estado completo de desnudez ó bien revestido exteriormente de
una capa de mielina, y pudiendo formar parte del sistema nervioso central ó del
sistema nervioso periférico.
Veamos
ahora cuáles son las relaciones de los diversos neurones en los centros
nerviosos.
Comunicaciones de los Neurones
Tanto
las fibras como las células nerviosas deben ser consideradas no como elementos
libres e independientes sino, por el contrario, como elementos que se hallan
recíprocamente unidos para constituir ese todo continuo, ese todo indivisible,
esa unidad nerviosa denominada neurón.
En
cuanto á la estructura de los centros nerviosos podemos decir que nos hallamos
en un nuevo día, puesto que la fórmula anteriormente dada, de que el sistema
nervioso se compone de elementos esenciales, como ser las fibras y las células
nerviosas, podernos substituirla por una expresión mucho más simple y al mismo
tiempo más exacta: «que el sistema nervioso no es más que el conjunto de un
gran número de neurones.»
Estos
neurones
se
diferencian muchísimo entre sí, no solo por lo que se refiere á su
significación fisiológica sino también a su disposición anatómica, variando
para cada uno de ellos según sea su función.
Es
pues necesario, para poder luego interpretar mejor los fenómenos complejos en
los cuales el eje cerebroespinal es el asiento, conocer muy bien las
relaciones recíprocas de los diferentes neurones.
Esta
cuestión de importancia capital ha permanecido durante mucho tiempo sumida en
la más completa oscuridad: pero en estos últimos años gracias a la aplicación
de diversos métodos técnicos, ha podido ser resuelta de una manera más ó menos
satisfactoria.
A
este objeto examinaremos sucesivamente las concepciones antiguas y las
concepciones nuevas que se tienen acerca del neurón.
Concepciones
antiguas
Las
concepciones que denominamos antiguas se basan en este hecho anatómico
(completamente hipotético, puesto que nunca -se ha comprobado), que
las células nerviosas se anastomosan entre sí en el espesor de la sustancia
gris.
Este
hecho ha inspirado un sinnúmero de teorías de las cuales dos se consideran como
esenciales y se las conoce con el nombre de los histólogos que las concibieron:
como ser Gerlach y Golgi; de ahí sus nombres de «Teoría de Gerlach» y «Teoría
de Golgi».
Teoría de
Gerlach
En
1871 Gerlach emite la primera teoría de conjunto acerca de la agrupación
recíproca de los elementos nerviosos.
Según
Gerlach, las prolongaciones protoplasmáticas de las células nerviosas,
juntamente con sus divisiones y subdivisiones, se transforman sucesivamente en
un gran número de fibrillas muy finas y delicadas que se anastomosan entre
ellas primeramente, y luego con las prolongaciones similares de las células
nerviosas vecinas. De este modo resulta la formación de una red difusa y
continua en toda la extensión de la sustancia gris, para cuya formación
concurren toda las prolongaciones protoplasmáticas de las células nerviosas.
Esta
red denominada «red de Gerlach» sirve para ligar las células nerviosas que las
constituyen, y así se explican las acciones diversas que ejercen unas células
sobre otras, ya sea en el estado fisiológico normal como en el estado
patológico.
Pero
esto no es todo: sobre cierto punto de la red en cuestión se ve surgir una
cierta cantidad de fibrillas que se dirigen todas a un punto común: allí
convergen, y, al unirse, forman un corto tronco nervioso que algo más lejos se
rodea de mielina y toma el aspecto de un cilindro-eje.
Existen,
pues, según la concepción de Gerlach, dos clases de cilindro-ejes, ó lo que es
lo mismo dos órdenes de fibras nerviosas: unas, las fibras ordinarias, que son
universalmente admitidas y que provienen, por la prolongación de Deiters, de la
célula nerviosa misma; y otras, que tendrían su origen en la red de Gerlach, y
por lo tanto, emanarían, como la misma red, de las prolongaciones dendríticas.
En
el trabajo de Gerlach por primera vez se expresa formalmente esta idea de red nerviosa continua
que,
a simple vista, parece lógica; de ahí que en un principio fue considerada como
un postulatum necesario.
En
efecto, esta idea de continuidad en las unidades superiores del sistema
nervioso es muy seductora: puesto que es más fácil concebirla en esta forma que
en aquella que supone que un órgano caracterizado por la unidad de acción esté compuesto de una serie de elementos
discontinuos.
Es
por esto que se explica, sin duda,
el gran numero de partidarios que ha tenido la teoría de una red continua: sin embargo, es una teoría sin base,
simplemente ideada para satisfacer el espíritu científico, pues está fundada
en el examen de cortes tratados por el cloruro de oro, de cortes coloreados al
carmín y luego disociados.
Estos
métodos no permiten comprobar en la sustancia gris otra cosa que un desorden
completo de prolongaciones celulares, pero en aquella época esto bastaba y la
idea de que el sistema nervioso estuviera compuesto de elementos discontinuos
hubiera sido absurda.
La
red ínterprotoplasmática de Gerlach no ha resistido á las observaciones hechas
por parte de Max Schultze y de Heule con ayuda de otros métodos técnicos: de
ahí que hoy día no despierte sino un simple interés histórico.
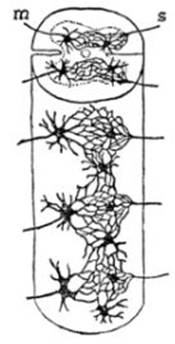
FIG. 1 – Esquema de la teoría de Gerlach
según Cajal. La figura representa un trozo de la
médula espinal: la sustancia gris, cuyos límites están determinados por líneas
punteadas, está ocupada por una red difusa formada por las prolongaciones
dentríticas de todas las células nerviosas; á esta red llegan las fibras de las
raíces sensitivas S; de las células
del cuerno anterior de la sustancia gris parten en forma
de cilindro-ejes las fibras de las raíces motrices m. Para completar este esquema hay que suponer que parten de la red
difusa fibras que se dirigen hacia el cerebro para llevar a ese órgano las impresiones
sensitivas y que, por otra parte, salen de esa misma red fibras que vienen del cerebro para llevar a la
médula las órdenes de los movimientos.
Teoría de Golgi
Poco
tiempo después, en 1873, Camilo Golgi hizo un descubrimiento que debía
revolucionar la histología del sistema nervioso.
Habiendo
sumergido Golgi en una solución de nitrato de plata fragmentos de tejido
nervioso fijados por bicromato de potasa, vio que ciertas células nerviosas se
coloreaban de negro y que al mismo tiempo se producía un depósito del mismo
color en la solución. Este depósito invadía las más finas expansiones de las células,
alterando así el color amarillo claro del fondo del corte y hacía resaltar esos
elementos – ofreciendo pues, ellos, un aspecto hasta entonces desconocido.
De
los métodos de coloración y de disociación, hasta entonces empleados, ninguno
había permitido comprobar la riqueza y la extensión considerable de las
arborizaciones que emanan de las células nerviosas.
La
Fig. N° 3 representa una preparación de León Azoulay, una célula nerviosa de la
corteza cerebral del hombre coloreada por el método de Golgi: se ve claramente
cuán rica es la célula en prolongaciones protoplasmáticas ó dendríticas, cuyas
ramas están erizadas de espinas ó apéndices periformes (más tarde nos ocuparemos
de la función que, se suponía, efectuaban esas espinas.)
De
la base de la célula se ve partir el cilindro-eje, prolongación más bien
delgada, de calibre uniforme, que emite finas ramitas colaterales: como este
cilindro-eje es el encargado de trasmitir las excitaciones motrices voluntarias
a la médula espinal, se comprende que su extensión será considerable comparada
con el volumen [sic] del cuerpo
celular.
Comparando
ahora esta figura con aquellas que se obtenían antes con la ayuda de la disociación
y la coloración del carmín, se comprenderá cuál habrá sido la sorpresa y
perplejidad de los histologistas cuando apareció el método de Golgi.
Pero
esta sorpresa se desvaneció cuando Pablo Ehrlich en 1886 descubrió el método
de coloración de los tejidos nerviosos por el azul de metileno, el cual daba
los mismos resultados que el método de Golgi; de ahí pues que fué necesario
admitir que la impregnación del cromato de plata responde á la realidad ó, al
menos, muy aproximadamente.
La
importancia de este método está en que si bien es cierto que no da más que
siluetas, éstas sin embargo, son exactas; por esta razón, histologistas, en
su mayoría alemanes, se han sublevado contra este método «unilateral», contra
esta técnica que no permite el conocimiento del contenido de las células. A
esto puede responderse que más de una técnica es «unilateral», es decir, que se
propone el estudio de una sola parte y no del conjunto para hacerlo de una manera
más precisa: además el estudio de un punto tan complejo como lo es sistema
nervioso no puede hacerse de otro modo que por la colaboración de un gran
número de métodos.
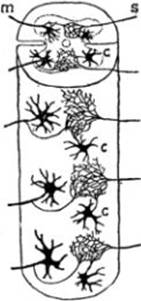
FIG. 2 – Esquema de la teoría de Golgi
según Ramón y Cajal; S, fibras de las
raíces sensitivas; m, fibra de las
raíces motrices; CC células del tipo
II cuyos cilindro-ejes terminan en arborizaciones muy ricas; del anastómosis de
las ramificaciones de esos cilindros-ejes con las raíces posteriores y el de
las colaterales de los cilindro-ejes de las células de los cuernos anteriores
resulta una red de cilindro-ejes difusa.
Indudablemente,
el desarrollo científico en esta materia se hizo de una manera lógica; si el
método de Golgi tuvo durante cierto tiempo tal preponderancia que conmoviera a
algunos, fue porque antes de estudiar los detalles del interior de las células
nerviosas; debía desmezclarse la forma de sus elementos y sus límites
interiores.
En
efecto, todos los descubrimientos objetivos hechos por Golgi son de suma importancia:
marcan en la histología del sistema nervioso un progreso capital.
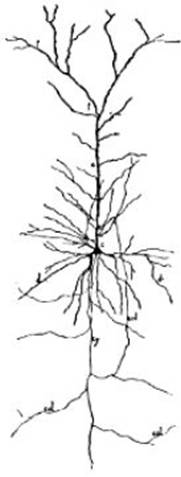
FIG. 3 – Célula de la corteza cerebral
del hombre (célula del tipo I de Golgi coloreada por el método de Golgi, dibujo
de Azoulay, según sus preparaciones). c, cuerpo celular; a, rama
protoplasmática; pp, ramificaciones protoplasmáticas ó dendríticas
formando penachos; dd, dendríticas de la base; cy, cilindro eje; col, colaterales del cilindro-eje. Véase
las espinas que erizan las dendríticas.
Golgi
puso de manifiesto los tres hechos siguientes:
1°
Las expansiones protoplasmáticas no se anastomosan entre si; se terminan libremente.
Esto
importa la negación completa de la red Gerlach.
2°
Todas las células nerviosas, sin excepción, poseen un cilindro eje.
3°
Los cilindro-ejes están ramificados y por consiguiente hay que distinguir, en
las células que las originan, tipos.
a)
Células en las que el cilindro-eje largo
no emite más que finas ramas colaterales, contribuyendo á formar la
sustancia blanca: estas células se denominan células del tipo 1° de Golgi.
b) Células en que el cilindro-eje corto se transforma a poca distancia de
la célula en ramificaciones sucesivas, formando una rica arborización destinada
a permanecer en una región poco extendida de la sustancia gris que le rodea:
estas células se denominan células del tipo II de Golgi; figura N° 4.
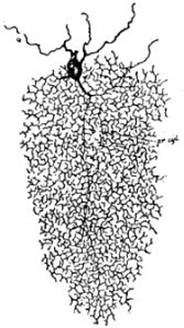
FIG. 4 – Célula de cilindro-eje corto
(célula del tipo II de Golgi) de la corteza del cerebelo de un gatito, coloreada
por el método de Golgi, según Van Gehuchten: prolongación cilindro-eje pr
cyl. Las ramificaciones de esa prolongación forman una arborización de una
riqueza extrema.
Estas conclusiones son hechos positivos de gran
valor, pero Golgi, al querer
agruparlas en una teoría de estructura general, no pudo separarse del ambiente
que le rodeaba: la red de Gerlach que acababa de ser demolida fue de nuevo
reconstruida con la ayuda de los elementos descubiertos: los cilindro-ejes
cortos y los colaterales de los cilindro-ejes largos. Supone Golgi que esas ramificaciones
se anastomosan para constituir una red nerviosa continua como la de Gerlach,
pero formadas por prolongaciones cilindro-ejes y no protoplasmáticas.
Como
vemos, la teoría de Golgi presenta gran analogía con la teoría de Gerlach, pues
en ambas es una red anastomótica lo que une a las células entre sí, y no
podía ser de otra manera pues la idea de
continuidad necesaria de la red nerviosa estaba arraigada profundamente
en el espíritu de los hombres científicos de aquella época.
Su
diferenciación consiste en que para Gerlach esta red estaría formada
exclusivamente por las prolongaciones protoplasmáticas, mientras que para
Golgi por las fibrillas desprendidas de las prolongaciones cilindro-ejes:
siendo una red interprotoplasmática en el primer caso e intercilindro-eje en el
segundo. Esta teoría, como la de Gerlach, está actualmente abandonada.
Concepciones
nuevas
En
1888, Ramón y Cajal substituía el procedimiento lento de coloración empleado
por Golgi por el procedimiento rápido que se denomina procedimiento de la doble
ó triple impregnación.
Después de perfeccionar el método, utilizando preferentemente
los sujetos jóvenes lo aplica sucesivamente al estudio de la médula, del
cerebro, del bulbo olfatorio, de los centros ópticos, del gran simpático, etc.
Con
la ayuda de este método pudo ver que no
existe ni la red de Golg¡ ni la de Gerlach; descubre que las
extremidades de los cilindro-ejes son libres, estableciendo así la
unidad anatómica del neurón.
Según
Cajal, las extremidades libres se ponen en comunicación por contigüidad; no
con prolongaciones similares, sino con prolongaciones protoplasmáticas ó con
el cuerpo mismo de otras células nerviosas (figura numero 5).
De
estudios hechos de los diversos órganos nerviosos y, especialmente, del cerebro
y retina llega Cajal á deducir que la corriente
nerviosa es centrífuga en los cilindro-ejes, y pasando esta corriente nerviosa
de neurón a neurón, se efectúa del cilindro-eje a las prolongaciones
protoplasmáticas o al cuerpo celular.
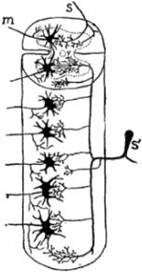
FIG. 5 – Esquema de la teoría del neurón
según Ramón y Cajal. S, S', fibra de
las raíces sensitivas y célula de los ganglios raquídeos de donde las fibras de
ésta categoría emanan; la célula S'
es unipolar, pero su única prolongación se bifurca inmediatamente y dá origen á
una fibra que se dirige a la periferia (nervio periférico sensitivo) y a una
fibra que se dirige hacia la médula (raíz posterior ó sensitiva). Cuando llega
a la médula la fibra radicular se bifurca, y dá una rama ascendente y una rama
descendente: cada una de esas ramas dan aún colaterales que se ponen en
comunicación por “contigüidad” con las células de la médula. Para mayor simplicidad
se ha imaginado que las células motrices de donde parten las fibras de las raíces
motrices son la m como lo indica la figura.
Esto
es lo que constituye «la teoría de la polarización dinámica de los elementos
nerviosos», por la que el neurón aparece como verdadera unidad fisiológica
(figura número 6).
Cajal
confirma también las ideas de His sobre la evolución de la célula y del
cilindro-eje: demuestra claramente cómo el cilindro-eje embrionario tiene en
su extremidad un abultamiento semejante a una yema que se conoce con el
nombre de cono de crecimiento y en donde tienen lugar los fenómenos nutritivos
por los cuales el cilindroeje se extiende más y más: de este modo precisa la
unidad embriológica del neurón.
Para
poder comprender bien el valor de la demostración anatómica de Cajal, es necesario
considerar las diversas conexiones de las células de Purkinje, pues es este
estudio el que le dio la intuición de su teoría.
La
célula de Purkinje está situada en la capa superficial de la corteza cerebelosa;
su forma lo indican las figs. números 7 y 8.
Sabemos
que sus prolongaciones protoplasmáticas están dispuestas en un solo plano,
estando todas orientadas en el mismo sentido: todas las ramitas de estas
prolongaciones se terminan libremente y en su trayecto presentan gran número de
espinas colaterales.
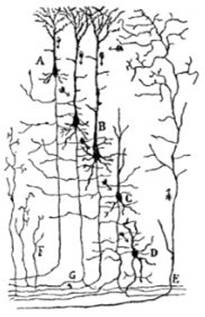
FIG. 6 – Figura semi-esquemática que
indica la marcha probable de las corrientes y las conexiones entre los
cilindro-ejes y ramificaciones protoplasmáticas en la corteza cerebral, según
Ramón y Cajal; A, pequeña célula piramidal; B, célula grande
piramidal; C y D, corpúsculos polimorfos; E, fibra terminal que viene de otros
centros; F, colateral de la sustancia blanca; G, cilindro-eje bifurcado en la sustancia blanca.
El
cilindro-eje desciende a la sustancia blanca y emite colaterales
ascendentes, los cuales tocan en varios puntos a las ramitas de las otras
células de Purkinje, viniendo pues a constituir entre esos elementos una cierta
solidaridad de acción: es esto,
pues, una conexión, si bien, es cierto,
una de las menos importantes de las muchas que sostiene dicha célula.
Próximas
á esta célula existen otras células estrelladas cuyo cilindro-eje sube
horizontalmente en la corteza y emite numerosos colaterales descendentes.
Cajal
pudo observar que esos colaterales al terminar, se ramifican y logran así
formar cestillos ó canastos terminales,
los que abrazan fuertemente el cuerpo
celular de la célula de Purkinje sin que por esto se confunda: el conjunto de estas fibrillas
se condensa en la
parte
inferior del cuerpo celular y forma así una especie de pincel, que envuelve la
primera parte del cilindro-eje de la célula de Purkinje.
Por
la aplicación de un nuevo método, Cajal llega a colorear las más finas expansiones
de las células nerviosas denominadas neuro-fibrillas y aparecen las relaciones
que sostienen dichas fibrillas con las células nerviosas: estas relaciones desconocidas
hasta entonces las observa Cajal en la célula de Purkinje: luego encontramos
una segunda manera ó modo de conexión.

FIG. 7 – Célula de Purkinje del cerebelo
humano: método de Golgi según Ramón y Cajal; a cilindro-eje; b
colateral ascendente; c y d arborizaciones protoplasmáticas.
Observa
también Cajal que sobre la capa de las células de Purkinje existen células
redondeadas pequeñísimas á las que se les conoce con el nombre de granos del cerebro: estas células tienen
prolongaciones protoplasmáticas cortas, en número de 3 ó 4; su prolongación cilindro-eje
asciende hacia la zona molecular y se divide en "7" y viene así a
constituir una fibra perpendicular a las arborizaciones de las células de
Purkinje; carecen de ramitas colaterales y van a terminarse en una espesura:
estas fibras, constituyen indudablemente, las vías de unión entre los granos
del cerebelo y las células de Purkinje.
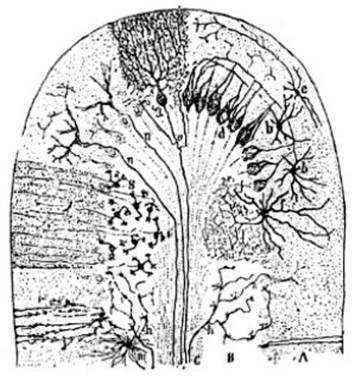
FIG. 8 – Corte transversal
semi-esquemático de una circunvolución cerebelosa de mamífero. A, zona molecular;
B, zona de granos; C, zona de la sustancia blanca; a célula de Purkinje
vista de frente; b pequeñas células estrelladas de la zona molecular
(células de cestillos terminales); d arborizaciones terminales
descendentes que rodean á la célula de Purkinje (cestillos terminales); e
células estrelladas superficiales; f grandes células estrelladas de la
capa de granos (células de cilindro-ejes cortos ó células del tipo II de
Golgi); g granos con sus cilindro-ejes bifurcados en i; h
fibras mohosas; n fibras trepadoras; o colaterales ascendentes
del cilindro-eje de la célula de Purkinje; j m células neuróticas
(tejido de sostenimiento) según Ramón y Cajal.
Cajal
encontró, aún, otra manera ó modo de conexión en la célula de Purkinje; en
efecto, en la sustancia blanca existen fibras nerviosas recubiertas con
mielina, cuya procedencia no se ha determinado todavía, las que se aplican á
las ramitas nerviosas de las células de Purkinje; se les denomina fibras
trepadoras, y terminan por medio de arborizaciones que se adhieren á los troncos
primarios ó secundarios de los corpúsculos de Purkinje.
Todas
estas conexiones pueden ser estudiadas indistintamente por el método de Golgi,
de Ehrlich, Ramón y Cajal, etc., y es de estas observaciones que el
histologista de Barcelona llega a las siguientes conclusiones:
1°
«Las arborizaciones protoplasmáticas y el cuerpo de las células tienen por
función, igual a los cilindro-ejes, la de transmitir las corrientes nerviosas,
puesto que hemos visto a ciertos cilindro-ejes ramificarse y terminar libremente
sobre las expansiones protoplasmáticas.»
2°
«Una célula nerviosa puede sostener conexiones independientes las unas de las
otras con diversos elementos, pues como hemos podido notarlo en las células de
Purkinje, el cuerpo celular se pone en comunicación por medio de los cestillos ó canastos terminales con los corpúsculos
estrellados de la capa molecular: la rama principal y las otras ascendentes se
ponen en comunicación por medio de las ramas trepadoras con los elementos de
la médula ó del cerebro y los ramúsculos protoplasmáticos terminales se ponen
en contacto con los cilindro-ejes de los granos ó fibras paralelas.»
3°
«La conexión dinámica de las células nerviosas parece que tiene lugar por el
contacto entre las arborizaciones finales de los cilindro-ejes, por una parte,
y por el cuerpo ó las ramificaciones protoplasmáticas de los corpúsculos
ganglionares por otra parte.»
Estas
conexiones que se verifican en las células de Purkinje se observan en todo el
sistema nervioso, como lo han demostrado por el método de Golgi, de Cox, de
Erhlich, Cajal y sus discípulos, y en general, observadores infatigables como Held, His, Kölliker,
Waldeyer, Azoulay, Lenhossek, Retzius, Van Gehuchten, etc.
Podemos,
pues, afirmar que la teoría del neurón tiene un valor general para el conjunto
del sistema nervioso y puede ser aplicable á todos los animales, con la restricción de
que ciertos invertebrados no han podido ser estudiados aún con los métodos
técnicos con que se conocen.
Criticas acerca
de esta teoría
Esta
teoría, como las demás, ha sido objeto de críticas y actualmente sigue
siéndolo: era de esperarse, pues todo descubrimiento trae consigo, desde el
momento en que se le conoce, partidarios y adversarios que, sugestionados por
las bondades ó errores de la teoría, llegan a caer en extremos completamente
falsos.
Efectivamente,
mientras unos no ven la anatomía, la psicología y la patología del sistema
nervioso más que al través del neurón, otros no solamente consideran falsa a esta teoría,
sino que van aún más lejos: la consideran como perjudicial, puesto que según
ellos tiene el defecto de detener todo progreso en el estudio del sistema
nervioso: su satisfacción solamente sería completa si llegasen a desterrar, de
la anatomía del sistema nervioso, el nombre de neurón.
Como
se ve, hay indudablemente en los dos sentidos exageración, lo cual vamos demostrar.
Hemos
visto que la teoría del neurón es hoy día una teoría anatómica; si hay
independencia anatómica la teoría es exacta; si hay continuidad entre sus
elementos es falsa.
Para
encarar debidamente la cuestión, vamos a considerar esta teoría desde el punto
de vista anatómico, fisiológico y embriológico.
Las
primeras objeciones referentes a las comunicaciones de las prolongaciones
celulares propiamente dichas pueden ser divididas en dos órdenes: las antiguas
y las nuevas.
Objeciones
antiguas
Desde
que apareció la teoría de Ramón y Cajal, el histologista ruso Dogiel se esforzó
en demostrar que en todos los puntos en donde Ramón y Cajal sostenía que
existían juxtaposiciones existían anastomosis.
En
efecto, Dogiel sostuvo que por el método de Ehrlich podía poner en
evidencia gran número de anastomosis en la retina.
Más
tarde otros histólogos, al querer comprobar estos hechos, vieron que si existían
anastomosis éstas eran muy raras y cada cual lo interpretaba a su manera.
Mas
no por esto Ramón y Cajal se desalienta; por el contrario, por sus estudios y
observaciones continuas llega a la conclusión de que tales anastomosis son artificiales
y debidas solamente á la exposición al aire que el método de Ehrlich requiere
para las preparaciones.
De
ahí que estas objeciones estén ya completamente abandonadas.
Objeciones
nuevas
Estas
objeciones están basadas en un descubrimiento reciente: el de las mazas terminales.
Estas
fueron vistas en 1896 por Auerbach, pero luego fueron estudiadas por Held y Bethe, y aún
por Ramón y Cajal que fue el que les dio dicho nombre.
Este
descubrimiento es importantísimo, pues nos indica las terminaciones
pericelulares en el animal adulto, es decir, la terminación de los
cilindro-ejes destinados a poner en comunicación un neurón con otro: ha
demostrado, pues, que las relaciones sostenidas entre los neurones son mucho
más íntimas que lo que se suponía por el estudio de los animales en el estado
embrionario.
Veamos
ahora las objeciones que sostiene Held.
Held considera a los
pequeños órganos que recubren en gran cantidad el cuerpo celular y las
prolongaciones protoplasmáticas y que reciben las últimas ramificaciones de
los cilindro-ejes como el conjunto de todas las partículas elementales y
las denominadas neurosomos: los describe como un conjunto estrellado, completamente ligados los
unos a los otros por medio de finas anastomosis quedando así constituida una
red pericelular que se halla convenientemente aplicada en toda la extensión del
neurón a excepción del cilindro-eje que se termina en las mazas terminales.
Es
por medio de esta red que Held explica las relaciones entro los cilindro-ejes
de las células nerviosas, relaciones completamente libres.
Held
sostiene que esa misma red comunica exactamente las células nerviosas con las
prolongaciones cilindro-ejes que van a reunirse con ellas.
Pero
esto no es todo: hay aún una red externa que
vendría a desempeñar un función protectora; esto es, hay una red externa de Golgi; cada malla de esta
red contendría una maza terminal y
las comunicaciones entre las mazas terminales se haría pasando sobre las
fibras de la red de Golgi.
Tal
es la teoría de Held.
Donaggio
había ya visto las mismas figuras con ayuda de su método, pero las interpretó
de otra manera: pues pone en duda que la naturaleza de la red formada por las
mazas terminales sea
nerviosa.
Bethe la interpretó a su vez de otro modo: para él la red de Golgi es de
naturaleza nerviosa, pues supone que en ella van a terminar los cilindro-ejes y
cree que es por intermedio de esa red que las neurofibrillas pasan de un neurón a otro.
Veamos
ahora los resultados a que ha llegado Cajal.
Este
histologista empleando un nuevo método para la coloración de las neurofibrillas
pudo colorear también las mazas terminales; las figuras 9 y 10 reproducen
fielmente lo que ofrecen las preparaciones; de su observación se deduce el
error de la teoría de Held y de Bethe, puesto que no existe ninguna anastómosis
entre esas mazas.
Estas
mazas terminales as¡ como las colaterales se aplican convenientemente sobre la
membrana envolvente de las células y de sus prolongaciones protoplasmáticas,
pero respetando a los cilindro-ejes.
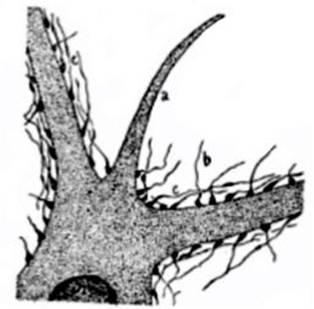
FIG. 9 – Porción de célula motriz de la médula
de un conejo adulto en donde se encuentran las mazas terminales; por el método
de plata reducida de Ramón y Cajal (dibujo según Ramón y Cajal); el protoplasma
celular ha sido representado arbitrariamente por puntitos y el núcleo n por puntitos
más juntos. Habiéndose hecho la preparación solamente en el centro de la
célula, las mazas terminales no se ven más que de perfil. Además de las mazas
terminales se ven las mazas laterales, es decir, colocadas al lado de una
fibra que continúa su trayecto ulteriormente; a cilindro-eje que no lleva
mazas mas que en su cono do origen; b fibras nerviosas convergentes; c fibras
nerviosas paralelas ó fibras del nido nervioso. El aumento con que esta célula
está representada puede ser avaluado en cerca de 1200 diámetros; se deduce que
cada maza terminal está representada como si tuviera un volumen aproximadamente
2 mil veces mayor que la realidad.
Su
estructura, generalmente alterable [ya] cuando las soluciones fijadoras son
muy débiles, explica el porqué las descripciones de Held, de Bethe y de
otros muchos sean equivocadas.
Held empleó el
método de Bethe, para el que se necesita hacer pasajes continuos de las piezas
en preparación, en soluciones ácidas y alcalinas que pueden alterar muchísimo
las estructuras finas; de ahí lo erróneo de sus conclusiones.
En
cambio Cajal usó fijadores excelentes tal como el nitrato de plata, de tal modo
que las preparaciones por él obtenidas pueden considerarse como exactas: de la
observación de estas por fuerza dedujo que no hay continuidad de sustancia, puesto
que cuando se colorean las mazas, se colorea también la célula permaneciendo
invisibles las fibrillas y cuando éstas se colorean, las células y las mazas
terminales se hacen invisibles.

FIG. 10 – Mazas terminales de la oliva
superior de un conejo adulto, por el método de plata reducida de Cajal; dibujo
según Ramón y Cajal. Tres células nerviosas, de las cuales una, la de la
derecha [izquierda del lector. MS], es vista por su superficie; el plano
dibujado pasa por el centro de otras células, lo que hace qué sea visto su
núcleo, y que, por el contrario, las mazas terminales no aparezcan sino en su periferia;
se ven también las ramificaciones protoplasmáticas de las células vecinas.
Esta
cuestión no tiene aún una solución completa; se apoya en conjeturas sucediendo
lo mismo con la ya referida red anastomática pericelular cuya existencia no se
halla definitivamente demostrada.
Hemos
dicho que la teoría del neurón podía ser considerada para su examen crítico desde el punto de vista fisiológico.
Esta
parte es de una importancia secundaria porque solamente está basada en
simples deducciones.
Puede
resumirse así: estando el neurón aislado constituye indudablemente una unidad
fisiológica y sus conexiones diversas demuestran que la corriente nerviosa es
centrípeta en las prolongaciones protoplasmáticas y centrífuga en las cilindro-ejes.
De
tal modo presentada la fisiología del sistema nervioso y en
particular del neurón, parece aceptable; sin embargo, veremos más tarde que
ha sido atacada por razones completamente teóricas por parte de Bethe.
Los
partidarios de la teoría del neurón han llegado al extremo, como hemos dicho
antes, de explicar todo por esa teoría: en efecto, Lépine y Matías Duval han
querido explicar el pasaje de la actividad al reposo por movimientos
amiboidales de las prolongaciones nerviosas, por medio de los cuales se
acercaban éstas ó se alejaban según el estado fisiológico.
Esta
teoría se apoyaba en una sola comprobación positiva: la de los movimientos
amiboidales vistos por Widertheim en las células del cerebro de un crustáceo
transparente, la Leptodora hyalina; pero
esta comprobación no fue después verificada.
Una
teoría semejante, lo que es un caso curioso, fue emitida por Rückardt cuando
aún se admitía la continuidad de la red nerviosa: la única diferencia es que
éste admitía la formación de rupturas en esa red durante el período inactivo.
Esta
teoría del sueño tuvo gran boga y fue la que inspiró gran número de trabajos:
algunos experimentadores llegaron hasta sostener que existían ciertas modificaciones
en las espinas de las prolongaciones protoplasmáticas cuando se erizaban y
otros como Azoulay han llegado a determinar que en realidad tienen lugar
alteraciones patológicas en dichas espinas.
Estas
hipótesis no resisten hoy a la crítica, puesto que las relaciones que existen
entre el cuerpo celular y las prolongaciones diversas que emanan de ella son
mucho más íntimas de lo que se creía.
Como
vemos, estas hipótesis están basadas en deducciones; de ahí el que no tengan
gran valor; lo que se puede decir, es que estas teorías permiten interpretar
los hechos fisiológicos conocidos; pero en la actualidad ninguna necesidad
fisiológica nos obliga a aceptarlas.
No
sabemos lo que es el influjo nervioso; de modo pues, que podemos suponer que
circula en una red continua, ó que se trasmite de célula a célula por el contacto
íntimo que ya hemos descrito ó, por último, que se propaga por una serie de
inducciones en una cadena discontinua: Todo esto puede ser verosímil.
Antes era fácil la explicación de las
continuaciones ó interrupciones de las corrientes nerviosas; pera con el
descubrimiento de las mazas terminales se ha obscurecido esta explicación.
Las
hipótesis acerca de las propiedades fisiológicas del neurón nos llevan a
considerar también sus propiedades patológicas.
Los partidarios de la teoría del neurón llegan a
sostener que la anatomía patológica no se puede comprender sin la ayuda de la
teoría del neurón.
Como veremos, esto es una afirmación exagerada.
Efectivamente, del examen de hechos anátomo-patológicos
resulta que existe individualidad celular en los órganos del sistema nervioso
como en los demás órganos, mas ninguno de esos hechos prueban que esa individualidad
sea absoluta al punto de negar toda anastómosis.
Lo
exacto es que el término neurón sirve para expresar
más bien que para explicar todas estas reacciones patológicas, lo que no quiere
decir que su uso sea indispensable.
Es
evidente que la hipótesis del neurón sirve para precisar la zona de degeneración en una región
determinada ó localizada del sistema nervioso: pero no siempre los límites son
los de los neurones
atacados,
pues es necesario tener en cuenta las degeneraciones secundarias y terciarias y
que existen en realidad y que obran indirectamente sobre diversos neurones que están ligados
entre sí con los neurones atacados: no es, pues, necesario llegar al extremo
de afirmar la individualidad absoluta de estos órganos ó elementos nerviosos
porque los mismos hechos pueden ser explicados siendo la individualidad
relativa.
Supuesta
la poca valía de estos asuntos, será más conveniente separarlos de la teoría
del neurón desde que no hacen más que recargarla sin reportarle el mínimo elemento
de demostración.
Por
el contrario, la embriología ofrece a la teoría del neurón cimientos muy
sólidos.
Bethe
ha renovado una teoría muy interesante; la de la estructura catenaria o cadenaria del cilindro eje imaginada
ya por Schwann.
Veamos
en qué consiste: el cilindro eje nace, según Bethe, independientemente de la
célula y no se une a ella sino algún tiempo después; está formado por las
células de los segmentos interanulares que elaboran cada cual un trozo; los
diversos trozos así formados se yuxtaponen y forman el cilindro-eje.
El
neurón, según esto, es un complexo celular.
Esta
idea es antigua: fue aceptada por los embriologistas que empleaban para
estudiar sus diversas piezas las técnicas que no permiten ver el cilindro-eje
sino los primeros bosquejos de los nervios en forma de hileras longitudinales
en la célula.
En
el estado adulto, el cilindro-eje ofrece un encogimiento a cada estrechez
ínteranular y ese aspecto robustece aún más la teoría cadenaria ó catenaria.
Agrega
Bethe, como hecho confirmante de su teoría, la siguiente experiencia que él
efectuó en varios animales jóvenes: corta un trozo del nervio ciático y ve al
poco tiempo que el trozo periférico queda aislado del trozo central y que se va
llenando de tubos nerviosos jóvenes; esos tubos nerviosos, según él, se han
desarrollado sobre esta parte por regeneración autógena sin la intervención de
una célula nerviosa.
Se
ha encontrado también en diversas investigaciones la existencia de nervios
motores bien desarrollados en monstruos [anencéfalos. MS] privados de sus centros
nerviosos.
Estos
dos argumentos si no fueran atacables serían suficientes para sostener la
teoría de Bethe: pero no es así.
El
primer argumento reproduce la antigua opinión de Philippeaux y Vulpian
sostenida por éstos en 1853, llegando ambos, después de gran número de
experiencias, a conclusiones semejantes a las de Bethe.
El
mismo Vulpian en 1874 reconoce los errores de su antigua teoría por lo que
desmiente sus primeras afirmaciones.
Recientemente
Munzer de Praga criticó acaloradamente la teoría de Bethe y por
experiencias análogas que efectuó llegó a conclusiones distintas.
En
el último Congreso Internacional de Psicología J. N Langley y H. K. Anderson igualmente han
criticado la teoría de Bethe, no reconociendo como verídica la generación
autógena de los nervios.
No
por esto Bethe deja de sostener su teoría, pues supone que no han de ser del
todo absurdas sus suposiciones cuando Van Gehuchten llega a los mismos resultados
que él.
A
los argumentos de Bethe, ha respondido Cajal anticipadamente con el
descubrimiento de los conos de
crecimiento de los cilindro-ejes.
Aunque
ya he hablado de este descubrimiento me parece lógico hacer aquí una
descripción breve de ellos para poder así comprender su importancia
considerable en el funcionamiento de los elementos nerviosos.
Empleando
uno de los métodos electivos se puede asistir, o, mejor dicho, seguir al cilindro-eje
en las diversas fases de su evolución; y podremos de tal suerte afirmar que
esta simple expansión celular se desarrolla progresivamente hacia la periferia
creciendo del mismo modo que una raíz ó la rama de un árbol.
En
los pollos, a los 3 ó 4 días de incubación, los cilindros-ejes de las raíces
anteriores, de las raíces posteriores y de la comisura anterior de la médula
se terminan muy cerca de su origen por una extremidad libre a cuyos lados se
notan mazas más ó menos abultadas de forma cónica, las cuales están provistas
de alas ó apéndices terminales que formarán más tarde los colaterales.
Según
que el embrión sea más ó menos adelantado puede verse que los cilindro-ejes
brotan, llegándose así a reconstituir sobre una serie de piezas las fases
sucesivas de su crecimiento.
R.
G. Harrison, cirujano muy hábil, hizo en el instituto Bonn investigaciones
interesantísimas sobre el desarrollo de los nervios y ha confirmado
completamente las afirmaciones de His y de Cajal.
Encontró
primero en las aletas de la cola de las larvas del tritón, con una longitud de
10 micrones, los conos de crecimiento en la extremidad de los cilindro-ejes sensitivos;
los describe como condensaciones con seudópodos y por el aspecto de estos conos
de crecimiento le surge el mismo pensamiento que a Cajal, es decir que el
crecimiento de esas fibras es el resultado de la actividad amiboidal del protoplasma en su extremidad.
Además,
ha comprobado que estas fibras sensitivas están en un comienzo desnudas, esto
es cuando se desarrollan; las células de Schwan, ó sean las células destinadas
a formar la vaina de mielina y que en la hipótesis de Bethe formarán en otras
el cilindro-eje por trozos, nacen de ganglios y se extienden sobre las partes
periféricas fibras que quedan algún tiempo desnudas.
Finalmente, por una experiencia muy ingeniosa,
suprimiendo por medio de un tijeretazo en embriones de ranas la parte
existente nerviosa destinada a producir los ganglios, Harrison pudo obtener el
desarrollo de los nervios motores faltando las células de Schwan; y logró
transmitir el influjo nervioso a los músculos, de donde se desprende que el
desarrollo del cilindro-eje no tiene nada que ver con las células de Schwan
(que provienen en su mayoría del ganglio raquídeo ó de las regiones
vecinas) como se suponía antes.
_______
VIAS NERVIOSAS: NEURONES Y NEUROFIBRILLAS — II
SUMARIO
-Qué se
entiende por neurofibrillas
-Teorías
antiguas
-Max Schultze
-Ranvier
-Nissl
-Held
-Teorías
de Apathy: sus trabajos en los invertebrados
De
los hechos enumerados en el capítulo anterior se deduce que, actualmente, la
concepción histológica del neurón no puede ser atacada ni rechazada, siempre
que se la considere dentro de los límites que le han fijado los técnicos.
La
sustancia nerviosa colorada por el método de Golgi ó de Ehrlich llega
precisamente hasta el límite determinado por Ramón y Cajal, sin continuarse en
la sustancia similar nerviosa de las células vecinas.
Pero
surge un problema, planteado recientemente y cuya solución es, por lo tanto,
objeto de dudas.
En
efecto, nos preguntamos ¿no hay otra sustancia nerviosa cuyos filamentos, no siendo colorables
por el método de Golgi, se extiendan fuera del límite del neurón de una célula
a otra, sin solución de continuidad? Y existiendo esos filamentos, ¿no desempeñarán
una función capital en la fisiología nerviosa?
Esta
hipótesis de las fibrillas independientes no es del todo inverosímil. Apathy y Bethe afirman
que las células se comunican entre sí por medio de neurofibrillas, las que ha
logrado colorar para su comprobación.
La
existencia de tales fibras en el interior de las células nerviosas es para los histologistas conocida de tan antiguo como las células mismas.
En
1844 Remack afirmaba su existencia y, en 1871, Max Schultze daba una
descripción de conjunto de dichas fibras, siendo considerada exacta
actualmente por Bethe: Ranvier, también, las ha estudiado.
Pero
el estudio preciso de ellas comenzó a hacerse después de los descubrimientos
técnicos de Apathy
y
luego de Bethe. Estos dos autores, por medio de numerosas experiencias, sostienen
que existe una red ó trayecto
extracelular ó intercelular, el cual llena la laguna dejada por el método de Golgi.
Establecen así una comunicación por continuidad directa entre los diversos neurones.
Posteriormente
Donaggio, Simarro y Ramón y Cajal han ideado técnicas nuevas para colorar las
neurofibrillas; pero, sobre todo, Ramón y Cajal ha hecho recientemente trabajos
notabilísimos sobre esta cuestión, que han proporcionado argumentos de primer
orden en contra de la existencia
de la porción extracelular de las redes de Apatthy y Bethe.
Con
esto, por consiguiente, se ha logrado reforzar la teoría del neurón, combatida
a tal punto que se llegó a pensar en su «decadencia».
Pero,
antes de estudiar estos trabajos de actualidad, será conveniente considerar
rápidamente las opiniones de los antepasados.
Teorías
antiguas
Max
Schultze sostiene que la célula nerviosa está formada por fibrillas sumergidas
en una sustancia interfibrilar granulosa: estas fibrillas son excesivamente
finas y atraviesan la células sin ramificarse ni anastomosarse, pasando ya de
una prolongación protoplasmática a otra, ó bien, de una prolongación protoplasmática
a un cilindroeje.
En
una prolongación celular, las fibrillas se reúnen formando un haz que en un
comienzo es único, pero que luego se divide, dando haces de más en más pequeños
a las ramas nacidas por bifurcaciones sucesivas de esa prolongación; pero esas
últimas ramificaciones de las fibras nerviosas no contienen más que una sola
fibrilla.
Según
esta disposición, las células vienen a ser simples lazos de distribución y
entrecruzamiento de las fibrillas nerviosas; luego, su función es secundaria desde el punto de vista anatómico; el único elemento esencial es la
fibrilla, que tiene una independencia anatómica y fisiológica completa. Cada
fibrilla es, pues, una unidad que tiene una función precisa, que es la de transportar el influjo nervioso de un punto determinado a otro también
determinado.
En
cuanto a las diferentes fibrillas de una misma célula, pueden efectuar
diferentes funciones, siendo independientes en su funcionamiento las unas de
las otras.
Ranvier,
por su parte, sostiene que las células nerviosas poseen una capa cortical fibrilar y que
las prolongaciones ramificadas de las células están formadas de fibrillas, las
cuales prolongan las del cuerpo celular.
En
cuanto al cilindroeje, Ranvier afirma también que tiene una constitución
fibrilar, puesto que lo considera destinado a ramificarse ulteriormente y a
descomponerse en «fibrillas terminales».
Ranvier está conforme con
Max Schultze en lo que se refiere a la individualidad anatómica y fisiológica
de las fibrillas, pero en cuanto a la célula nerviosa, no.
En
efecto, Ranvier concede a la célula una importancia fisiológica tal, que lo
que para Max Schultze no era más que una simple encrucijada destinada al
entrecruzamiento de las fibrillas nerviosas, para él es un órgano central del
cual emana gran número de fibras nerviosas cuyo funcionamiento es
trascendental.
Sostiene además Ranvier que, en todo el trayecto
recorrido, las fibrillas no permanecen agrupadas en las prolongaciones de las
células de donde emanan, sino que han de relacionarse con fibrillas formadas
por fibras nerviosas que provienen de otras células.
Un
ejemplo que comprueba a esta concepción de Ranvier, son las fibrillas del
plexo de la córnea; estas fibrillas se unen a las vecinas, para formar nuevas
fibras nerviosas que contienen, cada una, fibrillas de diferentes orígenes
celulares.
Del
mismo modo que la teoría de Bethe no es más que la teoría de Max Schultze
rejuvenecida, la teoría de Apathy lo es de la teoría de Ranvier; pues fácilmente
podemos establecer un paralelo entre el plexo de la córnea, según Ranvier, y la
neuropilema de los ganglios de la sanguijuela, según el anatomista húngaro.
Mientras
Remak, Max Schultze y Ranvier formaban esta «teoría fibrilar» acerca de la
estructura de las células nerviosas, otros autores, Bütschli en particular,
Nissl y sus continuadores luego, trataban de demostrar que los diversos
aspectos que ofrecen las células nerviosas en su estructura responden a una «estructura
reticular» del protoplasma nervioso.
Autores
como Held sostienen que este
protoplasma es homogéneo y que las redes ó grillas que se ven en su interior no
son mas que simples coagulaciones formadas por la intensidad de los reactivos
empleados en las preparaciones.
Hemos
visto, ligeramente, todas las teorías emitidas sobre la estructura del
protoplasma en general; nos ocuparemos ahora, primeramente, del aspecto de
estas fibrillas y el papel que los autores modernos les asignan en las
comunicaciones intercelulares y, por lo tanto, en la fisiología nerviosa.
Los
autores de cuyas concepciones hemos hablado no han visto, por medio de los
medios de preparación empleados, más que una pequeña estriación en el interior
de las células, la cual les dá ese aspecto fibrilar; siendo esto debido a la
diferencia de refringencia de las substancias que componen esos elementos.
Stephan
Apathy, profesor de la Universidad de Koloswar, fue el primero que encontró un procedimiento
técnico que permite colorar electivamente las fibrillas, dejando sin color la
sustancia interfibrilar. Esto es lo que permite distinguir claramente aquello
que, antes de él, apenas se podía entrever.
Aplicando
este procedimiento técnico, Apathy ha podido estudiar el sistema nervioso de ciertos
gusanos, especialmente el de las sanguijuelas. No pudo aplicar convenientemente
sus técnicas para el estudio de los animales superiores.
Para
poder comprender sus descubrimientos, vamos a hacer algunas consideraciones
acerca del sistema nervioso de los gusanos.
Cuando
se abre una sanguijuela por el lado ventral se ve, aplicado al intestino, un hilo delgado
blanquecino, rectilíneo, que se extiende de un extremo a otro del animal; esté hilo presenta, de trecho en
trecho, nudos que no son más que pequeños ganglios redondeados, de donde
parten los nervios periféricos.
Las
porciones del hilo comprendidas entre dos nudos toman el nombre de conectivos;
un examen más detenido nos permite comprobar que el hilo es doble: luego hay
dos conectivos juntos uno al otro, entre dos ganglios consecutivos.
Con
un aumento poco considerable se ve que los ganglios están formados por una
corona de células unipolares, cuyas prolongaciones se sumergen en una sustancia
central denominada «sustancia punteada de Leydig».
Esta
sustancia punteada está formada por una red de esas prolongaciones y de sus
innumerables ramificaciones: de esta sustancia parten, primero, los conectivos
que unen el ganglio con el que le precede, para seguir luego con la cadena; segundo,
los nervios periféricos.
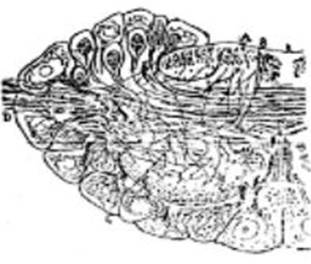
FIG: 11 – Corte transversal de un ganglio
nervioso de la sanguijuela, tratado por el método de Cajal. A, corte de fibras longitudinales
destinados a formar los conectivos que unen los ganglios entre sí; B, capa de cilindroejes comisurales ó
motores cruzados, es decir, cilindroejes provenientes de células situadas a un
lado del cuerpo y destinadas a ponerse en comunicación con células
ganglionares, ó con músculos situados del lado opuesto; C, plexo de neurofibrillas de la sustancia punteada de Leydig, ó
neuropilema ó red elemental de Apathy; D,
nervio periférico. Las células ganglionares unipolares forman una corona
alrededor de la sustancia punteada; las fibrillas de su red propia se continúan
con las de la red (Apathy) ó fieltradura (Cajal) de la sustancia punteada.
Las
células nerviosas de las sanguijuelas que no podían ser coloradas por el método
de Golgi, han sido estudiadas por Retzius con el azul de Ehrlich. El esquema N° 11 de
Cajal, que reproducimos, se refiere a los gusanos; pero, como el sistema
nervioso de las sanguijuelas tiene grandes analogías con el de los gusanos,
sin inconveniente, pues, podemos referir a las sanguijuelas las nociones generales
adquiridas acerca de los gusanos.
En
los gusanos de tierra, las células son piriformes y unipolares como en las
sanguijuelas; su extremidad más delgada se extiende en una sola fibra que
pronto se ramifica infinitamente.
Estas
ramificaciones, en su mayoría, terminan libremente en la sustancia punteada de
Leydig; sin
embargo,
una de ellas se extiende luego en los nervios periféricos para ir a ramificarse
en los músculos.
La
última ramificación es análoga a los cilindroejes de las células multipolares,
mientras que las primeras representan las prolongaciones protoplasmáticas: aparentemente
la última ramificación es una célula unipolar; pero no es así; por el
contrario, es una célula multipolar, cuyos polos se van acercando hasta confundirse
en uno solo a cierta distancia de la célula.
La Fig. N° 12
representa esquemáticamente las disposiciones que tienen los neurones en el sistema
nervioso de los gusanos.
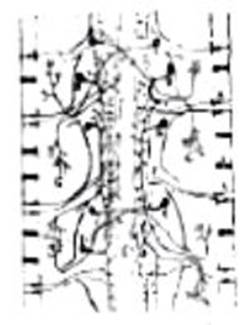
FIG. 12 – Esquema de la disposición
de los neurones en el sistema nervioso de los gusanos, según Cajal. A, células sensitivas de la piel; B, células motrices directas de los ganglios centrales; C, células motrices cruzadas; D, células
de asociación interganglionar; E, célula motriz multipolar; G, ramificaciones terminales de los
neurones motores en los músculos.
Las
impresiones del mundo exterior son transmitidas al sistema nervioso por fibras
que parten de las células bipolares situadas en la piel; éstas células de los
gusanos son semejantes a las células de los ganglios espinales de los animales
superiores.
Las
prolongaciones externas de las células son muy cortas, atraviesan la epidermis y se ponen así
directamente en comunicación con la superficie del cuerpo; las prolongaciones
internas tocan por intermedio de los nervios sensitivos a la sustancia punteada
de los ganglios donde se ramifican para ponerse en relación, por contacto, con
las ramificaciones protoplasmáticas de las células ganglionares.
Tal
es la estructura general del sistema nervioso en los gusanos, estudiada por los
métodos que permiten colorar esos elementos finísimos del sistema nervioso estando su
interpretación acorde con la teoría del neurón.
Nos
preguntamos ahora, ¿como se disponen las fibrillas nerviosas, descubiertas por Apathy, en el conjunto
del sistema nervioso?
Apathy
indudablemente
ha descubierto hechos importantísimos, cuyas interpretaciones son a menudo
discutidas; ha formulado una teoría estructural y fisiológica muy interesante,
puesto que trata de rebatir ciertas concepciones reinantes, pero de
consistencia efímera porque se basa en interpretaciones dudosas.
Los
hechos que pueden ser considerados hasta hoy definitivamente exactos, son: que
las células ganglionares y las células sensoriales contienen neurofibrillas de
tamaño variable que se dividen y anastomosan en redes diversamente
constituidas, según sean las células que se consideren; que se encuentran en el
cuerpo celular, casi siempre, dos redes, una periférica, la otra perinuclear,
que están unidas entre sí por fibras radiadas.
En
la sustancia punteada de Leydig existe una rica red de fibrillas
semejantes, que se hallan en continuidad con las fibrillas de las células
ganglionares; a esto es a lo que Apathy llama «red elemental difusa ó neuropilema»; y,
por último, que los filetes nerviosos contienen fibrillas de la misma
naturaleza que parten del neuropilema para ir a los músculos (nervios motores)
ó bien, que parten de las células sensoriales para ir al neuropilema (nervios
sensitivos).
La
existencia de las neurofibrillas es un hecho innegable; han sido vistas por Apathy por medio de
dos técnicas distintas; Bethe las ha observado empleando para ello su propio procedimiento
técnico y, finalmente, Cajal las ha estudiado claramente mediante su nuevo y
excelente método de plata reducida.
Por
medio de estos procederes técnicos se han obtenido representaciones
absolutamente iguales, operando en las sanguijuelas; pero no sucede lo mismo
cuando se opera en los vertebrados, porque el aspecto de las neurofibrillas
varía muchísimo según el método que se emplee.
Veamos
ligeramente la teoría general anátomo-fisiológica del sistema nervioso ideada
por Apathy
y
que se basa en estos hechos. Como las fibrillas son los elementos encargados
de conducir el fluido nervioso, viene de ahí que se les denomine «neurofibrillas»
ó «fibrillas conductrices». Las fibrillas que se observan en el neuropilema se
denominan fibrillas primitivas y se hallan formadas por la unión de fibrillas
elementales que se unen y desunen de infinitas maneras en las bifurcaciones y
en las redes.
Como
Apathy ha llegado a observar claramente la terminación de las fibrillas,
sostiene que estas forman una red interrumpida y difusa, no solamente en el
neuropilema de los ganglios, sino también en los músculos, en los órganos
sensoriales y, en general, en todas las partes del cuerpo.
Esa
red fibrilar se comunica con las células nerviosas de dos maneras, que son:
genética y funcional.
Apathy,
para
explicar esto, distingue en el sistema nervioso dos categorías de células: 1°
las «células nerviosas» que son las encargadas de elaborar las neurofibrillas
del mismo modo que las células musculares elaboran las fibrillas contráctiles ó
miofibrillas: 2° las «células ganglionares» que elaboran lo que debe ser conducido, es decir, el influjo nervioso;
una tercera clase de células debe agregarse a las células ganglionares y son
las «células sensoriales», que se asientan en la periferia.
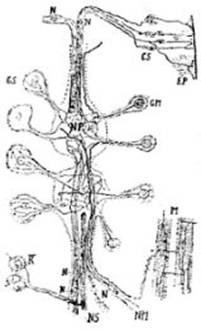
FIG. 13 – Esquema del sistema nervioso
en la sanguijuela, según Apathy. Ep, epidermis; Cs, las células sensoriales bipolares; R, dos células de la retina, cuyas
redes se comunican por anastomosis; N, células nerviosas, que son las células generadoras de las fibrillas; NS, nervio sensitivo, formado de fibrillas finas; NP, neuropilema, red elemental, en el
centro de cada ganglio, adonde llegan todas las fibrillas, sensitivas; GS, células ganglionares
sensitivas, en red
única de fibrillas finas; GM, células
ganglionares motrices,
cuya red periférica fina y la red perinuclear gruesa están reunidas por
fibrillas radiadas; NM,
nervio motor, conteniendo las gruesas fibrillas que emanan de la red perinuclear
y se colocan en el centro de la prolongación celular; M, músculo, con una red de neuroflbrillas
motrices no anastomosadas.
Compara
esta disposición con la de un telégrafo, en el cual los elementos de una
batería eléctrica se intercalan en el trayecto de un circuito de hilos, para
producir la energía destinada a ser transportada por esos hilos; y a la
circulación del fluido nervioso la compara con la circulación de la sangre.
La
figura núm. 13 representa esquemáticamente esta teoría.
De
la periferia, las fibrillas sensitivas finas después de haber atravesado las
células sensoriales van a terminar mediante los nervios sensitivos al
neuropilema de los ganglios: esas redes penetran, por una parte, en las
células ganglionares completamente sensitivas y, por otra parte, en las células
ganglionares motrices, para formar allí la fina red periférica; las fibrillas radiadas
de las células motrices pasan de la red fina periférica a la red perinuclear
más gruesa, de donde parten las voluminosas fibrillas motrices hacia los
músculos, para formar allí un plexo difuso, pero no anastomosado.
Toda
esta teoría se basa: primero, en una hipótesis que el mismo Apathy confiesa no
haberla verificado histológicamente, la cual consiste en el nacimiento de las
neurofibrillas dependiendo de las «células nerviosas»; segundo, sobre una
interpretación de imágenes microscópicas que es objeto de dudas; de ahí que sea
conveniente que la consideremos.
Apathy
ha
visto anastomosis entre las redes fibrilares en varios puntos de dos células
vecinas. Según él, las redes de las células de los ganglios de la cadena ganglionar se anastomosan
considerablemente por medio del neuropilema, que él considera como una red
anastomótica difusa y no como una fieltradura; por lo tanto existirían anastomosamientos
fibrilares directos entre las células ganglionares del plexo intestinal, entre
las células de la retina, entre ciertas células sensoriales de la piel, etc.
Ramón
y Cajal no ha visto anastomosis entre las células de los ganglios de la
cadena ganglionar; su método colorea claramente las neurofibrillas de las
sanguijuelas y
permite
efectuar cortes espesos, en los cuales es relativamente fácil poder seguir las
fibrillas
en
una gran extensión; para él, el neuropilema no es una «red anastomótica» sino
una simple fieltradura; ve terminaciones libres y bifurcaciones de las neurofibrillas
pero nunca anastomosis.
¿Cuál
de estos dos neurologistas tiene razón?
No
lo sabemos aún, el tiempo lo dirá ...
Con
el empleo del método de plata reducida no se ve ninguna anastomosis; sin
embargo, esas preparaciones son tan completas como las que obtiene Apathy en sus
trabajos: Azoulay no ha llegado a comprobar la existencia real de tales
anastomosis en las células.

FIG. 14 – Dos células del plexo intestinal
de una sanguijuela, según Apathy. Las líneas punteadas marcan los límites de
las células y de los nervios, así como el contorno del núcleo en cada una de
las dos células representadas, N° 1 y N° 2; M, fibra muscular a la que llega
una fibrilla nerviosa. Las células ganglionares, sus prolongaciones y los
nervios que de ella emanan están rodeados de una vaina común en la cual
solamente se colorean las fibrillas: es imposible, según Nageotte, distinguir
las neurofibrillas que pertenecen a una célula de las que pertenecen a una prolongación
proveniente de otra célula ó de un nervio que pasa junto a la primera.
La Fig. 14 reproduce la
complejidad extremada que existe en el interior de las células nerviosas; las
células y los nervios del plexo están contenidas en una vaina común y es imposible
determinar si una fibrilla penetra en una célula ó si está contenida en
algunas de las prolongaciones de la superficie.
Lo que puede ser
objeto de estudio fructífero son los ojos de la sanguijuela; por eso es que
nos ocuparemos de estudiar la disposición de sus neurofibrillas.
Ese
órgano está compuesto de
células sensoriales que son muy voluminosas y que están dispuestas en una sola
capa, la que se extiende alrededor de un eje ocupado por el nervio óptico, Fig. 15. Cada una de
ellas posee una prolongación muy fina que va al nervio; además, contienen un
vacuolo refringente (de forma irregular en la sanguijuela medicinal) limitado
por un borde de pestañas, las cuales vienen a desempeñar el papel de
cristalino.
De esas
células, las más profundas son pequeñas y no contienen más que un vacuolo rudimentario,
mientras que las más superficiales están dilatadas por un vacuolo tan grande
que aparecen como vesículas de paredes muy delgadas. El conjunto de este órgano
forma un cilindro colocado dentro de un estuche pigmentario.

FIG. 15 – Ojo de la sanguijuela visto
en un corte longitudinal; método de plata reducida; aumento de 200 diámetros. ep, epidermis; p, capa pigmentada coroides; n, n, nervio
óptico en el
ojo y fuera del ojo; c, p, células visuales profundas, pequeñas, de redes netamente
separadas; c, m, células visuales medias, colocadas las unas contra las otras; c,
s, células visuales superficiales, perforadas cada una por un grueso vacuolo, v. Las células visuales superficiales se adelantan en
realidad hasta abajo de la cutícula, y si aparecen aquí un poco distantes es
debido a una ligera inclinación del corte.
Las fibrillas se
disponen de modo que forman una red superficial muy rica en cada célula; y de
ella parte una neurofibrilla, que forma la única prolongación celular.
Apathy
sostiene
que existen anastomosis en las redes fibrilares de las células vecinas. Un
examen superficial las hace ostensibles, pero desaparecen ante un examen
profundo.
Las
redes de las diferentes células son totalmente independientes entre sí. En efecto,
en todos los puntos en que las disposiciones de las fibrillas son simples es
fácil comprobar que tales anastomosis no existen, v. g. las pequeñas células del fondo del ojo; en la mayoría de los
otros puntos pasa todo lo contrario: a cada paso se cree ver anastomosis, debido
a que las imágenes son muy obscuras y por lo tanto difíciles de analizar.
Debemos
tener en cuenta que las células retinianas son muy irregulares; tienen crestas
recortadas y onduladas, producidas por presiones recíprocas de las células;
además, las redes de las células vecinas están muy próximas y, como se hallan
situadas en la región superficial de la célula a que pertenecen, se forma una
fieltradura confusa que ofusca todo análisis, por ser las redes muy cerradas y
dispuestas en superficies irregulares.
Apathy
cree
que puede establecerse la existencia de las anastomosis por una serie de
dibujos, pero esto es dudoso; en efecto, es más difícil seguir una fibra en la
serie de planos de las diversas preparaciones que por un examen directo de
ellas, sobre todo si las imágenes son complicadas, como sucede a menudo.
Podemos
resolver esta cuestión disociando algunos cortes en una solución de
potasa al 40%, que no logra alterar la impregnación argéntica. Se verá que las
células se separan a su gusto, sin que dejen por esto rupturas en la red; de
ahí que podamos asegurar que en las redes fibrilares no existen anastomosis.
Por
lo tanto no existen las disposiciones de fibrillas en redes difusas, sostenidas
por Apathy y después por Bethe. La teoría de Apathy falla pues por su base; en
cambio parece tener razón Cajal, al asegurar que las redes fibrilares de los
diversos elementos celulares están aisladas unas de otra y no existen
anastomosis interneurónicos. _______
VIAS NERVIOSAS: NEURONES Y NEUROFIBRILLAS
– III
SUMARIO
-Teoría
de Bethe.
-Sus
trabajos en los vertebrados.
-Nuevos
métodos de Cajal.
-Teorías
nuevas.
-Funciones
que desempeñan las neurofibrillas, según Bethe y Cajal.
-Modernos
descubrimientos de Cajal y su discípulo Tello.
El
eminente profesor de la universidad de Estrasburgo, Albrech Bethe, encontró un
procedimiento técnico aplicable a los animales invertebrados solamente.
Por medio de este procedimiento pudo Bethe estudiar a estos
animales, principalmente a los crustáceos, y comprobar lo que ya había visto
Apathy, esto es, que el neuropilema no constituye una red difusa sino una serie
de departamentos distintos, cada uno de los cuales forma un centro funcional.
Además,
Bethe comprueba las ideas de Max Schultze de que en los vertebrados las
neurofibrillas pasan de una célula a otra. Tienen pues dos trayectos, uno
intracelular y otro extracelular.
Las
figuras núms. 16 y 17 corroboran las ideas de Bethe sobre las neurofibrillas
de los vertebrados, es decir, que las neurofibrillas pasan de una célula a otra
sin anastomosarse, de tal modo que la prolongación cilindroeje no se
diferencia en nada de la prolongación protoplasmática.
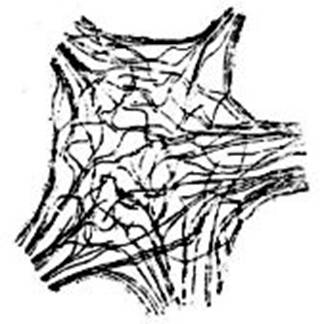
FIG. 16 – Células de los cuernos anteriores
de la médula; método y dibujo de Bethe. Las fibrillas pasan sin anastomosarse
de una prolongación á otra; algunas se bifurcan. Los bordes de la célula están
indicados por una línea punteada.
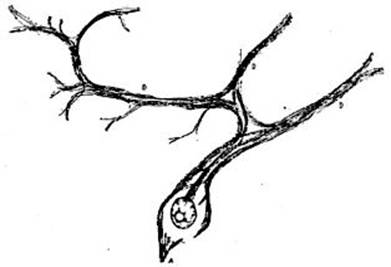
FIG. 17 – Célula de Purkinje del
cerebelo colorado por el método de Bethe; dibujo del mismo; A, cilindroeje; D,
D', D", ramificaciones dendríticas de diversas clases. Esta figura
muestra cómo las fibrillas pasan ya sea de una prolongación dendrítica al
cilindroeje, ó ya sea de una prolongación dendrítica a otra prolongación de la
misma naturaleza sin atravesar el cuerpo celular. Los límites de las células y
de sus prolongaciones están indicadas por una línea punteada.
Los
nuevos métodos de Cajal muestran con gran claridad que en las células de los
vertebrados existe una red fibrilar de extrema riqueza; red que varía según
sea la especie celular que se considere y que se halla formada por «fibrillas
primarias» que están a su vez reunidas por medio de fibrillas secundarias (figura
N° 18).
El
punto de mayor interés es el que se refiere a la existencia de las redes ó
trayectos extracelulares ó intercelulares de las fibrillas, afirmación que
importaría la
negación
completa del neurón. De ahí el que nos preguntemos si efectivamente existen
tales redes.
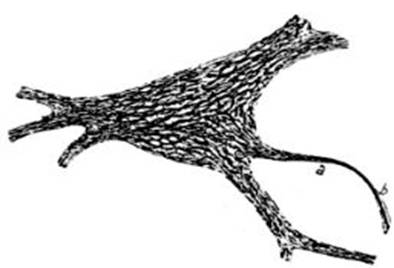
FIG. 18 – Célula motriz de la médula espinal
de un conejo de 15 días; método de Cajal de plata reducida; dibujo de Cajal. a, cilindroeje; b, punto en donde comienza la vaina de
mielina. Los límites del núcleo están indicados en el centro de la célula. La red de
neurofibrillas anastomosadas
forma mallas en donde se hallan los corpúsculos de Nissl; aquí no están colorados.
Bethe
afirma su existencia, pero Cajal la niega, sosteniendo que las neurofibrillas
permanecen estrictamente encerradas en los límites de los neurones.
Bethe
sostiene que las fibrillas pasan de un neurón a otro por medio de la red
externa de Golgi. Ya nos hemos ocupado de esta red entre cuyas mallas, según Held, se encuentran
agrupaciones estrelladas.
Estas
agrupaciones reciben, según Cajal, Held y Auerbach que las ha descubierto, las
extremidades de las ramificaciones cilindroejes aferentes.
Bethe
sostiene, además, que las extremidades cilindroejes terminan en la red de
Golgi. Las fibrillas del cilindroeje pasan a través de las bovedillas de esta
red y, como las fibrillas intracelulares penetran igualmente en las mismas
bovedillas, se llega a establecer allí una verdadera comunicación, entre las
fibrillas intracelulares y las fibrillas cilindroejes aferentes.
Las
bovedillas de la red externa de Golgi contienen, por lo tanto, la porción
extracelular de la red de las neurofibrillas.
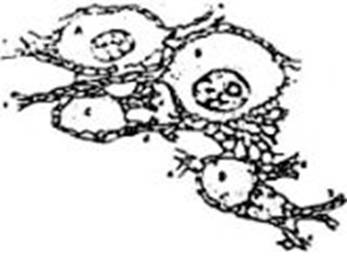
FIG. 19 – Cuatro células del núcleo
olivar del conejo; método y dibujo de Bethe. Red externa de Golgi en la superficie de las células A B C
D y de sus prolongaciones dendríticas d, d, d. Las redes de las células vecinas
se anastomosan entre sí.
Ahora
bien, nos preguntamos: ¿qué es esa red de Golgi? Indudablemente debemos
formularnos esta cuestión, porque el ilustre profesor de Padua, Golgi, ha
descubierto otra red, la «red interna» que está en la célula misma y parece
superponerse a la red canalicular é intracelular de Holmgreen.
Las
figuras núms. 19 y 20 muestran cómo esa red se extiende sobre la superficie de
las células y de sus prolongaciones; cómo pasa de una célula á otra,
constituyendo así una red difusa en toda la sustancia gris.
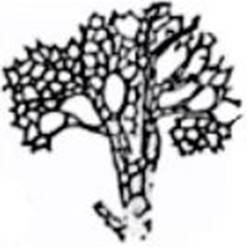
FIG. 20 – Ramificación terminal de una
prolongación protoplasmática de una célula del cuerno anterior de la médula de
buey sobre la superficie del cuerpo de otra célula de la misma naturaleza;
método y dibujo de Bethe. El protoplasma de la prolongación protoplasmática está
representada por una serie de puntos; el de la célula sobre la cual se terminan
las prolongaciones no se halla representada. La red de Golgi que rodea las ramificaciones de la
prolongación protoplasmática se continúa directamente con la de la célula a
la cual llega esa prolongación protoplasmática.
Golgi,
Donaggio, Bethe y Held
han
visto esta red y sostienen por lo tanto su existencia. Cajal cree que no se
trata en realidad de una red, sino de un simple coágulo nervioso.
Por
otra parte. los autores que sostienen la existencia de esa red le atribuyen
diferentes funciones: así, unos ven en ella un aparato de sostenimiento entre
las diversas células nerviosas, otros una simple red nerviosa, y Golgi vacila aún
acerca de estas interpretaciones.
Sin
embargo, Cajal y Held
sostienen
lo contrario de Betbe, puesto que demuestran claramente la terminación de los
cilindroejes en las otras formaciones nerviosas.
Adviértase
que si bien Bethe muestra gran confianza en sus ideas, al sostener la teoría de
las comunicaciones fibrilares por medio de redes va poco a poco perdiendo su seguridad,
cuando manifiesta que «algunas veces se puede ver el pasaje directo de las
ramificaciones cilindroejes en la red de Golgi, sobre
todo en la extremidad de las prolongaciones protoplasmáticas», y agrega que
«estos casos no son del todo demostrativos».
En
resumen, puede decirse que la técnica de Bethe está muy lejos de ser exacta y
que, por tanto, la comunicación de los neurones en los vertebrados, por medio
de neurofibrillas, descansa sobre una base falsa.
Completa
estas teorías modernas del neurón la concepción reciente de Franz Nissl.
Este
célebre histologista de Francfort, que ha descubierto y estudiado
minuciosamente la estructura de la «sustancia cromática» en las células
nerviosas, es un adversario encarnizado del neurón y ha publicado un volumen
inmenso en contra de la teoría del neurón, cuya lectura es muy fatigosa aún
hasta para los mismos alemanes.
Para
Nissl existe una sustancia: «el gris nervioso», que es la sustancia conductriz.
Esta sustancia se encuentra en el interior de las células, en donde forma
fibrillas, y existe también fuera de las células constituyendo gran parte de
la sustancia gris.
El
gran argumento de Nissl se funda en que existe entre las células un espacio
demasiado grande para que las redecillas de las prolongaciones celulares puedan
llenarlo. Después de una vasta discusión de 338 páginas el autor llega a esta
conclusión: «La existencia del gris nervioso, es decir, de una parte
constituida en su totalidad de sustancia nerviosa y no celular de la sustancia
gris, es una consecuencia, un hecho establecido, aunque la constitución histológica
es aún desconocida».
_______
Nos
hemos ocupado anatómicamente de las fibras nerviosas, así como de las más
recientes hipótesis fisiológicas que sobre ellas se tienen, pero no hemos
averiguado aún para qué sirven esas neurofibrillas, ni cuál es su importancia
funcional.
Examinemos,
pues, estos dos puntos.
La
fisiología de las células nerviosas es tan compleja como la de todas las otras
células del organismo; viven, es
decir, absorben, asimilan y excretan; funcionan
(función específica muy compleja) y, por último, resisten mecánicamente a las excitaciones exteriores.
¿Serán,
las fibrillas nerviosas, órganos de nutrición y de sostenimiento?
¿Serán
elementos encargados de la función específica?
En
este último caso, ¿cumplen toda la función nerviosa, ó tienen un papel
limitado, como ser el de conducir el fluido elaborado independientemente de
ellas? Estas cuestiones no pueden ser resueltas todavía, porque el número de observaciones
con que contamos es aún muy reducido.
Es
indudable que en los diversos tramos de la escala animal los órganos
encargados de la trasmisión del fluido nervioso, así como de su elaboración,
varían según sea su respectivo perfeccionamiento. En efecto, los organismos
unicelulares como las amibas tienen una función nerviosa rudimentaria: son
sensibles a la acción de las fuerzas exteriores, sufren a menudo las impresiones
recibidas, buscan las condiciones físicas más favorables a su mantenimiento y
se desplazan a fin de efectuar un hecho determinado.
En
los animales pluricelulares, la función nerviosa se halla desempeñada (en la
edad adulta, al menos) por células especiales.
No
se sabe aún si en los animales inferiores, como los celenterados, existen en
sus células nerviosas órganos (diferenciados) diversos; pero lo probable es
que en esos animales el protoplasma nervioso difiera sensiblemente del de los
animales superiores, puesto que todavía no se ha logrado colorarlos por los medios
técnicos electivos de Golgi y de Ehrlich.
Ya,
en los gusanos, las neurofibrillas han aparecido y en todos los demás animales
de organización superior existen, afectando disposiciones más complicadas a medida
que se va ascendiendo en la escala animal.
Esto
indica, pues, que las neurofibrillas se comportan como elementos fisiológicos
en la elaboración perfeccionada de los actos nerviosos complicados.
Las
neurofibrillas son una de las diversas estructuras del protoplasma nervioso; y
del mismo modo lo son el núcleo, órgano
excesivamente complejo que, según se supone, desempeña un papel importante en
el funcionamiento nervioso, puesto que sufre modificaciones cuando la célula
se halla debilitada por un trabajo prolongado; el jugo celular, porción transparente; un espongioplasma reticulado; la sustancia
cromática que se encuentra agrupada en las mallas del retículo.
A
estas formaciones podemos agregar la red
interna de Golgi que, probablemente, está superpuesta a la red de los canalículos de Holmgreen; y
la red externa de Golgi que,
según algunos autores, es un simple coágulo ó un aparato de sostén de los
elementos nerviosos, ó un medio de aislamiento ó una red que contiene la porción
extra-celular de las neurofibrillas en el pasaje de un neurón a otro.
Las
fibrillas nerviosas no son, pues, más que los rodajes de la máquina complicada
del elemento nervioso. Sin embargo, ese rodaje es de gran importancia en la
función nerviosa.
Efectivamente;
planteando la cuestión desde otro punto de vista, se puede comprobar que las
fibrillas no son únicamente órganos propios de las células nerviosas. Gran
número de células tienen formaciones análogas y, en todas ellas, esas fibrillas
desempeñan funciones muy importantes en la actividad celular.
Nos
bastará citar las fibrillas musculares, lisas y estriadas, las fibrillas de
las células de pestañas vibrátiles, las fibrillas de un haz conjuntivo y
tendinoso, etc.
En
todas estas formas celulares, las fibrillas se nos representan como productos
de una elaboración protoplasmática, destinadas ora á ser el instrumento del
trabajo específico de la célula que tiene movimiento, como la fibra muscular,
ora á una acción de sostenimiento como el tejido neuróglico.
De
ahí que, por analogía, podemos suponer que la fibrilla nerviosa no está
destinada a desempeñar funciones nutritivas, ni a consolidar a la célula en
sus funciones, sino que es, por el contrario, el órgano ó al menos uno de los
órganos principales de la función nerviosa.
Nos
preguntamos ahora ¿es sólo una parte ó bien toda la función nerviosa la que
desempeñan las neurofibrillas?
Según
Apathy,
el
acto nervioso se descompone en dos elementos:
1°
la elaboración del influjo nervioso por una especie de secreción efectuada por
el protoplasma de las células ganglionares;
2°
las trasmisiones de este fluido por las neurofibrillas,
Las
células ganglionares vienen así a constituir verdaderas pilas eléctricas, cuyo
circuito está formado por la red fibrilar.
A
primera vista, esta concepción es clara y nos seduce, pues se explica por la
noción experimental de que un fluido nervioso recorre los nervios con cierta
velocidad, circulando por los cilindroejes recubiertos exteriormente de
mielina: capa aisladora, como lo es la envoltura exterior de un cable. Empero,
esta teoría no reposa más que en la continuidad de la red fibrilar admitida por
Apathy.
Conociendo
como conocemos los elementos de convicción que ofrece la teoría de Apathy desde el punto
de vista anatómico, podemos afirmar que, hasta la fecha, ningún hecho
anatómico ó fisiológico ha probado que las neurofibrillas, ni la sustancia interfibrilar
como suponen algunos, sean las conductoras del fluido nervioso.
Bethe
tampoco establece distinciones entre la elaboración y la trasmisión del
influjo nervioso. Atribuye a las fibrillas la función nerviosa en «block» y por
las disposiciones anatómicas que él ha observado, deduce que el resto de la
célula efectúa únicamente funciones tróficas.
Para
sostener esta hipótesis, Bethe se basa en las experiencias que ha realizado en
los cangrejos. He aquí en qué consisten estas experiencias: cuando se toca la
duodécima antena de un cangrejo, se produce el movimiento reflejo de esta
antena; este movimiento tiene como centro un ganglio que se halla formado de un
neuropilema central y de células unipolares periféricas. La
disposición de estas células es tal que se les puede suprimir una gran parte
de su contenido sin que se llegue a tocar en lo mínimo al neuropilema. Después
de esta operación, el movimiento reflejo persiste durante dos días para luego
desaparecer.
Bethe
explica esta persistencia del movimiento reflejo admitiendo que la impresión
nerviosa es conducida del órgano sensorial al órgano motor por la red anastomótica
de las fibrillas del neuropilema; de donde deduce que, según toda probabilidad,
la función nerviosa pertenece a las neurofibrillas, pues las células
desempeñan funciones nutritivas.
Los
resultados de esta experiencia se comprenden fácilmente si se admite que el
neuropilema está constituido por un gran número de prolongaciones celulares que
contienen fibrillas no anastomosadas.
Sabemos,
en efecto, que las células unipolares poseen prolongaciones múltiples,
centrípetas unas, centrífugas otras, pero que nacen todas de un mismo tronco:
la ablación de los cuerpos celulares no modifica las conexiones de estas
prolongaciones; se puede, pues, admitir que, en la experiencia de Bethe, la
transmisión nerviosa se efectúa por la sustancia formada por las prolongaciones
celulares que han permanecido enteras y guardando relaciones normales; y no
solamente por
las
fibrillas.
En
realidad, esa experiencia sólo prueba que los neurones son capaces de
sobrevivir un tiempo limitado después de la ablación de una parte de la sustancia
que contiene el núcleo; y que durante este tiempo subsiste la función
nerviosa. Este fenómeno es semejante al que ofrecen los organismos celulares
que se cortan en trozos, en los que se observa que los fragmentos que no contienen
núcleos continúan viviendo y moviéndose durante un cierto tiempo, pasado el
cual mueren.
Si
la experiencia de Bethe no nos proporciona ninguna noción positiva acerca del
funcionamiento de las neurofibrillas, en cambio Cajal y su discípulo Tello han
logrado abrir un nuevo campo de investigaciones, con sus recientes
descubrimientos de fisiología nerviosa.
Estos
autores demuestran que la red de las neurofibrillas no es como se creía «constante», sino que es susceptible de
sufrir modificaciones más ó menos considerables según el estado de actividad
nerviosa en que se encuentre el animal en el momento de su muerte.
Fundan
sus conclusiones en experiencias realizadas en culebras y lagartos, es decir,
en animales sujetos al adormecimiento invernal fisiológico. Cajal ha hecho
también observaciones en animales atacados de rabia paralítica. Estas
experiencias consisten en lo siguiente
Cajal
y Tello observaron que durante el adormecimiento invernal de esos animales las
neurofibrillas de las células de la médula son poco numerosas y muy grandes (Fig. 21 A). Un día un
lagarto que se quebró la cola despertó de su sueño excitándose por este
accidente, y en las preparaciones que se siguieron Cajal observó, con sorpresa,
que las fibrillas gruesas y poco numerosas de antes se hallaban reemplazadas
por infinidad de fibrillas muy finas (Fig. 21 C).
Esta
observación fue para él un descubrimiento, porque pudo establecer que el
estado en que había observado al animal en un principio era el del
adormecimiento causado por el frío.
Comprobó
también que si se calienta al animal en una estufa, durante algunos instantes,
se despierta y agita vivamente; y que si se le mata en esos momentos se notan
en sus células fibrillas finas y muy numerosas.
Será
éste, pues, el aspecto de las células durante la estación caliente, cuando el
lagarto posee su actividad normal.
El estado
intermediario entre el adormecimiento y la actividad primaveral está
representado en la Fig. 21 B; se ve allí cómo las fibrillas gigantes se
disocian en fibrillas finas que se esparcen por toda la célula.
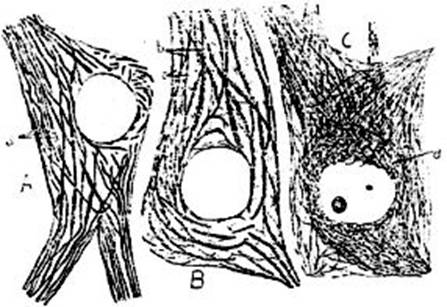
FIG. 21 – Grandes células nerviosas
intersticiales del bulbo del lagarto, según Tello; método de Cajal.
A.- Célula de un lagarto
adormecido por el frío.
Las fibrillas son enormes y poco numerosas, formadas en realidad por coalescencia
de fibrillas finas en haces; a, espesamientos sobre el trayecto de esos haces.
B.- Célula de un lagarto por
despertarse. Las
fibrillas finas están por esparcirse; b, haces formados por fibrillas finas que han comenzado a separarse
unas de otras; c, fibrillas
finas completamente desprendidas de los haces.
C.- Célula de un lagarto
despertado, en
plena actividad. Los haces no existen ya; todas las fibrillas finas están
esparcidas; d, red perinuclear.
Tales
son los hechos descubiertos por Cajal y Tello; son los primeros hechos
positivos que se refieren al mecanismo íntimo de la célula nerviosa.
Este
mecanismo no se ha explicado todavía, pero constituye una prueba evidente de
que las neurofibrillas efectúan funciones muy activas en el engranaje complejo
que se denomina sistema nervioso.
Hemos
pasado rápidamente en revista todas las concepciones que ha sugerido el
estudio de las vías nerviosas, falsas las unas y precisas las otras. Es de
esperarse que la precisión de esas concepciones no será duradera y que a ellas
seguirán nuevas observaciones y descubrimientos, que iluminen con luz más segura
los puntos que aún oscilan en torno de esa unidad histológica sumamente
complicada que se llama neurón.
MATILDE
V. VILLA,
Profesora
Normal.
_____________
Copyright
© 1997 Electroneurobiología. Este trabajo
original constituye un artículo de acceso público; su copia exacta y
redistribución por cualquier medio están permitidas bajo la condición de
conservar esta noticia y la referencia completa a su publicación incluyendo la
URL original (ver arriba). / This is an Open Access article: verbatim copying
and redistribution of this article are permitted in all media for any purpose,
provided this notice is preserved along with the article's full citation and
original URL (above).

revista
Electroneurobiología
ISSN: ONLINE 1850-1826 - PRINT 0328-0446

Haga doble
"click" en el título de cualquier artículo, para leerlo ahora -
Double-click on any article to read it now:
SOCIOLOGÍA DE LAS NEUROCIENCIAS
![]() Pour comprendre
l'enjeu : L’anthropologie ganglionnaire, un psychovirus démasqué (français)
Pour comprendre
l'enjeu : L’anthropologie ganglionnaire, un psychovirus démasqué (français)
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (190 kB:
recomendada) o .DOC (76 kB).
![]() L'aliénisme en
Argentine : Diego Alcorta (1827) : Dissertation sur la manie... aiguë?
(français)
L'aliénisme en
Argentine : Diego Alcorta (1827) : Dissertation sur la manie... aiguë?
(français)
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (600 kB: recomendada)
o .DOC (320 kB).
![]() Metaphors at odds in conceiving organismal-societal government: The Political Structure of the Brain: Cerebral Localization in Bismarckian Germany
(English)
Metaphors at odds in conceiving organismal-societal government: The Political Structure of the Brain: Cerebral Localization in Bismarckian Germany
(English)
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (228 kB: recomendada)
o .DOC (175 kB).
![]() One of the earliest recorded works in Biological Psychology presents a synthetic system of psychology weaved from positivist philosophy and the principles of physical chemistry
: The Biological Psychology of José Ingenieros, some biographical points, and Wilhelm Ostwald’s (Nobel Prize Chemistry, 1909) Introduction to the 1922 German edition
(English)
One of the earliest recorded works in Biological Psychology presents a synthetic system of psychology weaved from positivist philosophy and the principles of physical chemistry
: The Biological Psychology of José Ingenieros, some biographical points, and Wilhelm Ostwald’s (Nobel Prize Chemistry, 1909) Introduction to the 1922 German edition
(English)
Puede leer, imprimir o
guardar en su disco duro esta investigación en versión .PDF (990 kB: recomendada)
o .DOC (1258 kB).
en archivo .PDF (101 kB) o bien .DOC (84 kB)
ELECTRONEUROBIOLOGÍA
Efectos
relativísticos en biofísica cerebral:
Puede leer, imprimir o guardar en su disco duro esta investigación en versión .PDF (496 kB: recomendada) o .DOC (227 kB)
SUMARIO Y
PÁRRAFOS INICIALES EN CASTELLANO
Diversificación
de recursos electroneurobiológicos en la evolución del sistema nervioso:
Puede leer, imprimir o guardar en su disco duro esta investigación en versión .PDF (733 kB: recomendada) o .DOC (406 kB)
auch als .PDF (285 kB) oder .DOC (161 kB)
Puede leer, imprimir o guardar en su disco duro esta investigación
en versión .PDF (354 kB:
recomendada) o .DOC (236 kB).
Cálculo
de potenciales dentro de las células
 Calcule
intensidades eléctricas y magnéticas en cada compartimiento neuronal: The nervous principle: active versus passive electric
processes in neurons (Explains how to calculate electric and magnetic
field strengths inside different neuronal compartments) (LONG FILE IN ENGLISH with
Bulgarian, Russian and Spanish abstracts/TOCs)
Calcule
intensidades eléctricas y magnéticas en cada compartimiento neuronal: The nervous principle: active versus passive electric
processes in neurons (Explains how to calculate electric and magnetic
field strengths inside different neuronal compartments) (LONG FILE IN ENGLISH with
Bulgarian, Russian and Spanish abstracts/TOCs)
Podrá leer, imprimir o guardar en su disco duro esta investigación en versión .PDF (2 Mb): recomendada) o .DOC (1,5 Mb). También como .html comprimido (compressed .HTML folder: 0,5 Mb) .ZIP.
Evaluación de
potenciales fuera de las células
Signal analysis to exploit the information of steady-state recordings: Do’s and don’ts in Fourier analysis of steady-state potentials
(Assumptions in the discrete Fourier transform (DFT) not necessarily fulfilled in real-world applications) (English)
Minireview
The ‘Enabling’ Neural Correlates for All Consciousness: Latest Findings in the Mechanisms of Cortical ‘Arousal’
(Conditions in the neuronal and biochemical level skipping consideration of any more fundamental biophysical issues, e. g. quantum or relativistic) (English)
You can download a .PDF (269 kB:
recommended) or .DOC (128 kB)
file for printing this work




