Gobierno de la ciudad de Buenos Aires
Hospital Neuropsiqui�trico
"Dr. Jos� Tiburcio Borda"
Laboratorio de Investigaciones Electroneurobiol�gicas
y
Revista
Electroneurobiolog�a
ISSN: ONLINE 1850-1826 - PRINT 0328-0446

Los taxones mayores de la vida org�nica
y la nomenclatura de la vida en Marte:
primera clasificaci�n biol�gica de un
organismo marciano
�(ubicaci�n de
los agentes activos de la Misi�n Vikingo de 1976 en la taxonom�a y sistem�tica
biol�gica)
por
Mario Crocco
Director, Centro de Investigaciones�
Neurobiol�gicas, Ministerio de Salud, Rep�blica Argentina, y Jefe de,
Laboratorio de Investigaciones�
Electroneurobiol�gicas, Hospital "Dr. J. T. Borda", Gobierno
de la Ciudad de Buenos Aires
Ave. Amancio Alcorta 1602, 1283 Buenos Aires, Argentina; tel./fax (54)
11 4306 7314, mcrocco[at]electroneubio.com.ar
Electroneurobiolog�a 2007; 15 (2), pp. 1-34; URL <http://electroneubio.secyt.gov.ar/index2.htm>
Publication date / fecha de publicaci�n:� 14 de abril, 2007
seguido de una
Correcci�n: primera clasificaci�n biol�gica de un organismo marciano,
g�nero Gillevinia (no Levinia)
Correction note: first biological
classification of a Martian organism, genus Gillevinia
(not Levinia)
Electroneurobiolog�a 2007; 15 (2), pp. 35-37; URL
<http://electroneubio.secyt.gov.ar/index2.htm>
Publication date / fecha de publicaci�n:� 28 de abril, 2007
Copyright � April 2007 Electroneurobiolog�a. Este texto es un art�culo de
acceso p�blico; su copia exacta y redistribuci�n por cualquier medio est�n
permitidas bajo la condici�n de conservar esta noticia y la referencia completa
a su publicaci�n incluyendo la URL (ver arriba). / This is an Open Access
article: verbatim copying and redistribution of this article are permitted in
all media for any purpose, provided this notice is preserved along with the article's
full citation and URL (above).
Puede
obtener un archivo
.PDF
(recomendado: 1,3 MB) para leer o imprimir este art�culo, desde aqu� o de / You
can download a .PDF
(recommended: 1.3 MB) file for reading or printing, either from here or� <
http://electroneubio.secyt.gov.ar/index2.html >
La acci�n
nomenclatural objeto de la presente comunicaci�n fue ejecutada el 20 de
diciembre de 2006. El presente art�culo fue consecuentemente preparado en ingl�s
para su comunicaci�n en el exterior. Traducci�n de Mariela Szirko. Publicado el
14 de abril de 2007.
��
SUMARIO � Contexto: Debido
a su car�cter rudimentario, el presente estado de los conocimientos acerca de
los procesos de vida en Marte es propicio para establecer las principales
l�neas racionales destinadas a clasificar los dominios mayores de la vida
org�nica. Bajo las convenciones taxon�micas, una apropiada iniciativa
clasificatoria en esta etapa podr� despu�s prevenir complicaciones extracient�ficas
que afecten el an�lisis cient�fico de cualquier eventual variedad de formas
vivientes extraterrestres. Con ello como prop�sito, los espec�menes activos en
los experimentos de detecci�n de vida de la Mision Vikingo son nomenclados.
M�todo: Como precondici�n para este y cualquier prop�sito
similar, los dominios biol�gicos (o super-reinos) son agrupados en bi�sferas,
cuya denominaci�n formal � para establecerla sin alejarse del lenguaje natural
� se elige como la del cuerpo celeste continuo en que la agrupaci�n se
distribuye: bi�sfera terrestre, bi�sfera marciana, etc. Los nombres de las
bi�sferas se formar�n, pues, sobre el genitivo del nombre del pertinente cuerpo
continuo, planetario o satelital. Pero aun los nombres de las bi�sferas no
emergen como el �ltimo nivel nomenclatural requerido para una extensi�n consistente
de la ciencia biol�gica. Mientras que no existe necesidad absoluta de que todos
los cuerpos celestes biosfereados de hecho integren un sistema estelar, hay consenso
respecto a que la mayor�a de ellos lo hacen. Como, concebiblemente, las
agrupaciones de bi�sferas pueden compartir caracter�sticas relacionadas a su
estrella o ce�ido par binario central, el nombre de la estrella (el nombre tradicional
en la cultura del designador prioritario, quien elegir� el convencional de un
cat�logo estelar mayor cuando no se dispusiera de nombre tradicional para la
estrella o estrellas) puede ser pr�ctico para identificar los m�s grandes
sistemas de la vida org�nica. De tal modo, la agrupaci�n de las bi�sferas que
hubiera en nuestro sistema solar forma el sistema de vida org�nica Solaria.
Conclusi�n: El agente activo caracterizado por su
oportunamente comunicado comportamiento como responsable de los resultados del
experimento de liberaci�n marcada en la Misi�n Vikingo de 1976 formalmente
constituye, para la biolog�a sistem�tica, la siguiente entrada taxon�mica:
sistema de vida org�nica, Solaria; bi�sfera, Marciana; rei�no, Jakobia; genus et
species, Gillevinia* straata, todos ellos nova taxa.
* Nuevo nombre
de g�nero, en substituci�n de Levinia;
ver p�ginas 35-37
Palabras
clave: organismo vivo,
definici�n; vida org�nica, definici�n; ecolog�a; exobiolog�a; organismos extraterrestres;
taxonom�a y sistem�tica; bi�sfera de Marte; Gillevinia
straata
��
Abstract
Background: The current state of knowledge about life
processes in Mars, because of its rudimentariness, is propitious for setting up
the main rational lines aimed to classify organic life's major domains. Under
taxonomical conventions, a proper classificatory move at this time may later
prevent extrascientific complications in the scientific analysis of any
eventual variety of extraterrestrial life forms. With this in view, the active
specimens of the Viking Mission's Labeled Release life detection experiments
are nomenclated.
Method: As a precondition to this and any akin purpose,
biological domains (or superkingdoms) are clustered into biospheres, whose
formal name � so as to establish it without departing from natural language �
is chosen as that of the continuous astronomical body in which the cluster
distributes: terrestrial biosphere, martian biosphere, etc. Biosphere names are
thus formed upon the genitive of the pertinent planetary or satellital
continuous body's name. Still, they do not appear as the latest nomenclatural
level required for a consistent extension of biological science. While there is
no absolute need that all of the biosphered astronomical bodies actually belong
with a stellar system, there is by now a consensus that most of them do. As
clusters of biospheres might conceivably share features related to their central
star or tight binary pair, the name of the star (the traditional one within the
culture of the priority namer, who is to choose one from a major consensus
stellar catalog if no traditional name is available for the star or stars) may
become practical to identify life's grandest systems. Any cluster of biospheres
in our solar system thus form the organic life system Solaria.
Conclusion: The active agent characterized by its
reported behavior as responsible of the results of the 1976 Viking Mission's
Labeled Release experiment formally features, for systematic biology, the
following taxonomical entry: organic life system, Solaria; biosphere, Marciana;
kingdom, Jakobia; genus et species, Gillevinia
straata, all of them nova taxa as reported below.
Key words: living organism, definition; organic life, definition; ecology;
exobiology; extraterrestrial organisms; extraterrestrial taxa; taxonomy &
systematics; Mars biosphere; Gillevinia
straata
��
Introducci�n
El
presente estado del conocimiento humano sobre formas de vida extraterrestre es
inicial y por cierto desbalanceado. Comprende gran riqueza de datos te�ricos
provenientes de extrapolaciones de la biolog�a terrestre, ni una veintena de
resultados experimentales de sondeos metab�licos realizados sobre el planeta
Marte, y el conocido debate sobre restos microbianos en meteoritos provenientes
de ese planeta. Por ahora, eso es todo. El dif�cil acceso a otros cuerpos planetarios
torna lento, e inconexo, todo esfuerzo de espigar hechos referidos a otras
biosferas. Esa colecci�n inconexa y lenta ya fue observada antes, porque
tambi�n ocurri� en los comienzos de otras ciencias como la arqueolog�a moderna,
el diagn�stico por neuroim�genes, y algunas ramas de la f�sica muy dependientes
de los desarrollos tecnol�gicos. En la mayor parte de aquellos casos, el nuevo
territorio acad�mico comienza a delinearse como figuras at�picas, bastante dif�ciles
de interpretar. Gradualmente se hacen m�s claras y m�s f�ciles para obtener:
cabe pensar en las tempranas im�genes de difracci�n de compuestos org�nicos, o
sobre las nacientes ecografias y ventriculografias. A medida que los
principales elementos nuevos se hacen identificables, las etapas iniciales resultan
una oportunidad �nica para establecer la nomenclatura b�sica � lo que tambi�n
transporta ideas b�sicas o conceptos. Esto es uno de los puntos principales del
presente art�culo: algunas de tales ideas o conceptos facilitan y otras estorban
el progreso en el nuevo campo.
Uno de
estos conceptos b�sicos es la homogeneidad del dominio: las reservas sobre la
naturaleza homog�nea de un proceso o fen�meno pueden disuadir la colaboraci�n
plena, conteniendo el progreso, en base a escr�pulos no cient�ficos no raras
veces. Esto parece de especial importancia en el caso de extender la biolog�a
fuera de la Tierra � una tarea que se ha puesto en marcha durante nuestra vida.
Algunos
casos hist�ricos
La
nomenclatura es para esto un instrumento de valor. Lejos de ser s�lo materia de
catalogaci�n, la nomenclatura pone las fronteras as� como las articulaciones
ontol�gicas en el interior de un dominio acad�mico. Un caso hist�rico que
merece mencionarse es la substituci�n por Benjam�n Franklin de "electricidades"
m�s y menos en el lugar de las "electricidades" v�treas (positivas) y resinosas (negativas) de Carlos de
Cisternay Du Fay. Mientras en su tiempo la substituci�n de Franklin no import�
ning�n avance observacional e incluso, cabe decir, ning�n desarrollo conceptual,
y as� durante un siglo y m�s all� oper� para la mayor parte de los investigadores
como un mero cambio de nombres, la implicada homogeneidad "de los dos fl�idos"
contribuy� a presentar la electricidad como un solo bloque de fen�menos,
localizado ante un diferente bloque � presumiblemente tambi�n �nico, pues � de
fen�menos magn�ticos, simplificando y estimulando la b�squeda de sus relaciones
rec�procas, sobre las cuales se basa toda nuestra civilizaci�n tecnol�gica.
Otro
caso hist�rico pertinente, menos afortunado, es la unificaci�n por Arist�teles
de todas las entidades de inter�s biol�gico bajo la noci�n gen�rica de una
vida, zooe�, sostenida en todos los
casos por un alma o psyche� que puede
incluir experiencia o excluirla � y su vigente herencia de rompecabezas a menudo
verbales, legada a la psicolog�a y los consciousness
studies. Antes que Arist�teles creara la biolog�a, no se dispon�a de ning�n
tratamiento conceptual unificado para todos los organismos vivos conocidos. La
creaci�n de la biolog�a signific� el descubrimiento de un �nico hilo conceptual
para toda su variedad, lo que podr�a haberse alcanzado declarando que todos los
organismos vivos en cuanto vivos debieran estar dotados de experiencia, o bien
que en cuanto tan s�lo vivos debieran carecer de ella. Pero, como viene siendo
materia de creciente atenci�n en Historia de Ideas, Arist�teles ten�a serios motivos
para no distinguir entre la experiencia y el comportamiento no experienciado. A
causa de ello Arist�teles debi� escoger otro factor de unificaci�n: escogi� un
modelo tr�fico del conocimiento (informaci�n) y confusion� ambas categor�as,
las entidades experienciadoras y las no experimentantes, como organismos
vivientes basados en su conformaci�n din�mica, aportando as� serias incertidumbres
a la ontolog�a del psiquismo. Uno se siente tentado a establecer un paralelo
con los experimentos de detecci�n de vida de los informes publicados de la
Misi�n Vikingo, todav�a presentados como s�lo preliminares, y sus datos brutos
(Levin 2006a, p. 22: "los datos de la � Vikingo no han sido a�n [escrib�a
el 19 de noviembre de 2006] objetivamente revisados respecto a la evidencia de
vida"; p. 25, "un informe definitivo de los hallazgos y la conclusi�n
deber�a emitirse. A la fecha [el 19 de noviembre de 2006], no ha habido a�n ninguna
formal revisi�n de pares sobre este experimento y sus datos
relacionados"); y no puede menos de interrogarse si tal an�lisis incompleto
de los datos colectados tambi�n hubiera ocurrido en caso que los experimentos
se hubiesen referido a alg�n otro rinc�n inaccesible, pero terrestre.
Ampliaci�n
de los criterios de barrera biogeogr�fica
a las distancias
interplanetarias
�Cu�les
son pues, en cuanto a la nomenclatura biol�gica, los dominios principales de la
vida org�nica? El criterio de la sistem�tica filogen�tica, a saber la posici�n
fil�tica m�s o menos remota de un presunto antepasado com�n, es pr�cticamente
inaplicable y probablemente sea inadecuado para muchos dominios mayores de vida,
o para la mayor parte de pares de ellos. La clad�stica requiere una riqueza de
datos que no est� disponible durante las etapas iniciales de esta extensi�n
disciplinaria de la biolog�a m�s all� de la Tierra. Sin embargo, aunque de
ninguna manera el 'aislamiento' planetario sea absoluto, la dificultad misma en
el acceso, con la que nos topamos para cosechar conocimientos sobre la vida
extraterrestre, parece se�alar a la condici�n biogeogr�fica � la evoluci�n
discontinua de la vida sobre una pluralidad de cuerpos astron�micos continuos �
como el criterio m�s conveniente para distinguir esos dominios mayores de la vida
org�nica.
Las
bi�sferas, tanto si se las hallare florecientes en plena diversidad o en cambio
iniciales y rudimentarias al m�ximo, pueden reconocerse por la observaci�n de
un solo evento de la actividad propia de los organismos vivos, ya que cada uno
de estos eventos indica un linaje natural. Y debido a que, para cada dos
procesos biosf�ricos planetariamente aislados, se estima pr�cticamente
imposible la repetici�n razonablemente similar de etapas excepto las m�s
iniciales, la extensi�n de la biolog�a m�s all� de la Tierra parece bien
servida por la extensi�n, a distancias interplanetarias, de los criterios de
barrera biogeogr�fica. Esto sit�a a las bi�sferas
como los dominios principales de la vida para la nomenclatura biol�gica, y al
verbo biosferear como conveniente denominaci�n
para un curso posible de la evoluci�n f�sicoqu�mica planetaria o satelital.
Esto, a m�s, establece una clara barrera sistem�tica, incluso si pudiere
determinarse que algunas formas de vida iniciales o prebi�ticas comparten niveles
supraqu�micos de organizaci�n entre cuerpos
planetarios o satelitales.
Ser
dominios sistem�ticos principales de la vida no es lo mismo, sin embargo, que
ser sus taxones m�s amplios. Las bi�sferas no lo son. La misma extensi�n de
aquel criterio taxon�mico, de proveniencia biogeogr�fica, cuyo empleo primario
es distinguir poblaciones hasta el nivel de biosferas, encuentra adem�s un ensanchamiento
bastante natural en el sistema estelar
al que pertenece cualquier cuerpo astron�mico continuo que sostenga bi�sfera.
Esa referencia a una estrella central o prieto sistema binario exige, sin
embargo, una discusi�n breve, ya que en algunos casos tal referencia podr�a
quedar vac�a.
En la
historia de cualquier cuerpo astron�mico apropiado, sostener bi�sfera no puede
ser m�s que una condici�n transitoria. Y de seguro es concebible que una
peque�a proporci�n de planetas, o de cuerpos que antes fueran satelitales,
podr�an mantenerse todav�a biosfereados aun alg�n tiempo despu�s de haber sido
expulsados de su sistema estelar por alg�n acontecimiento astron�mico. En imaginables
circunstancias, orbitar alrededor de una estrella o prieto sistema binario
particular puede no ser necesario para conservar la bi�sfera. Por lo tanto, la
denominaci�n de los taxones m�s amplios de vida org�nica � a saber, de los sistemas de vida org�nica � en base a la
estrella, o prieto sistema binario estelar, que sostenga un planeta biosfereado
particular al que tal vez debieran a�adirse otros m�s, en casos posiblemente excepcionales
habr�a de hacer lugar para permanecer vac�a. Sin embargo, pienso que el
criterio deber�a ser conservado. Una raz�n es que, salvo tales excepciones
te�ricas, los cuerpos planetarios o satelitales demasiado alejados de la
radiaci�n estelar o ciclos de deformaci�n gravitatoria no parecen capaces de
sostener los flujos de energ�a necesarios para desarrollar una bi�sfera o para
mantenerla residualmente. Otra raz�n es que, al conectar formalmente los
taxones biol�gicos m�s amplios al desarrollo de cuerpos estelares, esto es a la
evoluci�n de bi�sferas separadas o racimos biosf�ricos desarrollados alrededor
de esos cuerpos, por intuici�n y conceptualmente se liga la biolog�a con la
evoluci�n astrof�sica, en una secuencia �nica de procesos de curso temporal: la
evoluci�n astrof�sico-biol�gica.
Circunstancias
de la presente acci�n nomenclatural
El
presente trabajo sobre la biolog�a sistem�tica de sistemas autoorganizativos es
ocasional en una tradici�n investigativa neurobiol�gica, de modo que resulta
apropiado perfilar las circunstancias en que surge. Desde 1914 nuestros
principales cient�ficos dedicaron especiales esfuerzos de investigaci�n a conceptuar
la vida, desde luego sin quitar el ojo � por tratarse de neurobi�logos � de la
distinci�n entre vida y psiquismo: tema este interesante (v�ase, por ejemplo,
Crocco 2007), seguramente ajeno al presente art�culo. En los tempranos a�os sesenta
me toc� el turno de ofrecer algunas definiciones, adoptadas a su debido tiempo,
y para fines de 2006 una revista t�cnica de esta repartici�n oficial [se trata
de Electroneurobiolog�a. N. d. T.],
revista dedicada a nuestros temas de investigaci�n y sobre la que tengo responsabilidad
administrativa, decidi� recopilar en una selecci�n especial varios art�culos antiguos
y recientes que estudian el concepto de vida. En noviembre de 2006 la directora
ejecutiva de la revista me trajo una remisi�n del Experimentador de los
experimentos de detecci�n de vida de la Misi�n Vikingo, Gilberto V. Levin. La
misma fue publicada el 30 de noviembre, hall�ndose disponible en versiones
inglesa (Levin 2006a) y castellana (Levin 2006b).
Notablemente,
tanto la cuesti�n sobre la que el trabajo informaba (un problema epistemol�gico
que afecta a los sondeos de vida que apunten a descubrir componentes biol�gicos
separados en vez de metabolismo) cuanto la noci�n de vida que manejaba eran
compatibles con el concepto de vida en nuestra tradici�n investigativa. Por
donde tres semanas m�s tarde, en una alocuci�n que form� parte del memorial por
el semicentenario del deceso de uno de nuestros cient�ficos (y Ministro de
Salud de la Naci�n), Ram�n Carrillo, quien en 1949 tambi�n trat� la cuesti�n de
la definici�n cient�fica de la vida, expuse el punto epistemol�gico de Levin,
evalu� seg�n lo mejor de nuestro conocimiento como metab�lica la evidencia
colectada en 1976, subray� como elemento nuevo el corriente foco
sobre simbiosis y din�mica endocitobiol�gica � propio de la biolog�a actual de
los organismos multicelulares � y el consiguiente encogimiento biol�gico, hoy corriente,
de las diferencias sist�micas entre organismos compuestos de endosimbiontes y,
por otra parte, biotas (como las que podr�an haber sido representadas en el
ensayo de las muestras de suelo marciano para sondear metabolismo), se�al� los
para entonces muy recientes informes sobre el hielo subsuperficial y el flujo
observado en las regueros
o arroyuelos en Marte (de
haber estado hablando despu�s de febrero, hubiera debido agregar los
indicadores de metano como un signo de actividad posible biol�gica y comentar
los escasos requerimientos de reparaci�n biol�gica planteados seg�n las nuevas
evaluaciones de rayos c�smicos en la superficie), y proporcion� al agente
activo en el suelo de Marte (bien sea semejante a una "especie"
bacterial terrestre � analizada en Cohan 2002 � o compuesto de especies como los hongos
liquenizantes, o bien a una muestra de
biota aun m�s diversificada) el
nombre cient�fico de Levinia
straata.
En las
semanas anteriores, la lectura acostumbrada de los resultados de Vikingo como
abi�ticos se hab�a recontextualizado r�pido, pasando Marte a ser m�s y m�s
reinterpretado como biosfereado. En octubre �ltimo, evaluando experimentalmente
la sensibilidad insuficiente de uno de los experimentos de los m�dulos de aterrizaje
para encontrar materia org�nica en el suelo, Navarro Gonz�lez et al. 2006 hab�a agregado mucho al
conjunto de pruebas que dejaron los resultados de Vikingo cada vez m�s abiertos
a lecturas biog�nicas. En noviembre y diciembre �ltimos salieron a luz las
observaciones de los flujos recientes de las regueros o arroyuelos (de un fl�ido a�n no identificado) y los
vol�menes enormes del hielo subsuperficial [cuyo volumen derretido cubrir�a todo
el planeta con una profundidad de once metros. N. d. T.]. El 30 de enero de
2007, la primera versi�n de Houtkooper y Schulze-Makuch 2007 intent� dar cuenta
de los resultados de la misi�n Vikingo suponi�ndoles un posible origen
biog�nico, como se comenta m�s abajo. Despu�s de esto en Buenos Aires, el dos
de febrero, la directora ejecutiva de nuestra revista public� un sumario
editorial en castellano (Szirko 2007), en el que coment� que, en vez de
intentar descubrir organismos vivos, las misiones futuras a Marte todav�a est�n
siendo planeadas para descubrir s�lo biomarcadores. El trabajo continuaba indicando
cu�n correcto es afirmar que la navaja de Ockham previene atribuir a organismos
vivos el hallazgo de cualquiera de sus componentes separados, creando as� una
situaci�n de tensi�n conceptual que constituye una seria cuesti�n epistemol�gica
para la investigaci�n de toda eventual bi�sfera marciana.
Consideraciones
breves
basadas en
la definici�n de vida
En
cuanto al mencionado concepto de vida adoptado en nuestra tradici�n, el punto aqu�
relevante consiste en que encontramos el l�mite, entre procesos de vida y
procesos no vivientes, en su referencia a una entidad externa � no en la composici�n
interior; y que definimos la vida org�nica como la convergencia de subprogramas que sostiene sistemas estoc�sticos
multivariados, esto es, como el
conjunto de procesos eficientemente causales dependientes de la organizaci�n de
dichos sistemas que, en ciertos escenarios o ambientes, le permiten a esa
organizaci�n continuar existiendo. Esto esclarece la fuerza de la objeci�n
epistemol�gica de Levin, la que � pienso � puede ser resumida advirtiendo, como
Szirko lo hace, que la navaja de Ockham previene atribuir a organismos vivos
todo descubrimiento de cualquiera de sus componentes separados.
De
donde resulta que la detecci�n confiable de vida no tiene nada que ver con las
concentraciones de compuestos org�nicos, perceptibles o no como marcaciones
moleculares con la sensitividad de alg�n instrumento, o por medio de la
separaci�n de amino�cidos de imagen especular uno del otro (estereois�meros).
Consideremos un poco m�s aquellos conceptos, para as� contextualizar m�s abajo
los argumentos acerca de la detecci�n de vida org�nica en la superficie de
Marte. Con ese mismo objetivo, saltar� aqu� sobre el contraste entre la convergencia
y la divergencia de los subprogramas interiores de las entidades biol�gicas,
contraste que opone los niveles sucesivos de integraci�n sist�mica (los que
pueden incluir una biocenosis o ecosistema, comunidad ecol�gica, o asociaciones
de tipo liquenizante; luego, cualquier organismo endocitobiologicamente
compuesto que resida all�; luego sus simbiontes, y sus reacciones bioqu�micas �
cada uno con su propio nivel de interacciones horizontales y verticales y de
selecci�n natural), porque este contraste, aunque fundamental, sin embargo no
se relaciona de inmediato con nuestro tema presente.
Que el
l�mite, entre procesos org�nicos vivientes y no vivientes, yazga en verdad en
su referencia a una entidad externa m�s bien que en la composici�n interior, se
hace manifiesto, por ejemplo, al pensar en virus cristalizados o tard�grados o
rot�feros crioconservados, cuya composici�n no var�a al recuperar la plenitud
de su actividad o "animaci�n". Tardigrados y bacterias congelados y
desecados, que son organismos, y as� tambi�n partes de organismos como la piel
de mam�feros, c�lulas de la c�rnea y hasta esperma liofilizado que todav�a
puede usarse para fertilizar �vulos de mam�fero cuando se lo reconstituye,
consisten en su totalidad de materia no quiral no viviente mientras est�n
congelados.
Sin
embargo, tras recuperar su solvente interno (agua en esos casos) y energ�a
termal, pueden comenzar de nuevo un conjunto
de procesos eficientemente causales dependientes de su propia organizaci�n que,
en ciertos escenarios o ambientes, le permiten a esa organizaci�n
continuar� existiendo. El metabolismo
del hospedador para el virus, y las condiciones de temperatura y humedad para
los m�s aut�nomos tard�grados, componen los escenarios ambientales que permiten
a su organizaci�n (molecular en el primer caso; cito- e histol�gica tambi�n, en
el segundo) continuar existiendo.
Respecto
al criterio de que la homoquiralidad, o cualquier predominio num�rico de los estereois�meros
con quiralidad levo o dextro sobre los de la otra, ser�a la prueba
"absoluta" de vida porque la evoluci�n habr�a encontrado demasiado complicado
el manejo de mezclas rac�micas (un criterio quiz�s tomado prestado de la idea
de que, en organismos complejos, se desarrollaron dos sexos m�s bien que m�s de
ellos debido a la supuestamente excesiva complejidad recombinacional de tres o
m�s sexos, n�mero que no obstante se encuentra y se supera largamente en
protozoos bastante complejos), tal criterio no puede distinguir "vida
f�sil" (semejante al combustible f�sil terrestre) de la vida existente.
As�, lejos de constituir ninguna prueba "absoluta" o siquiera
"robusta" de vida, el criterio de la homoquiralidad se muestra por s�
mismo del todo d�bil e incompleto, cayendo bajo las objeciones de principio
contra cualquier tentativa de descubrir la vida mediante el descubrimiento de
sus componentes separados. Ese criterio necesita as� complementarse con
escenarios ambientales espec�ficos para cada especie.
La
vida org�nica real, a saber la convergencia
de subprogramas que sostiene sistemas estoc�sticos multivariados, esto es, el conjunto de procesos eficientemente
causales dependientes de la organizaci�n de dichos sistemas que, en ciertos escenarios
o ambientes, le permiten a esa organizaci�n continuar existiendo, asume tales
escenarios ambientales espec�ficos para cada especie y puede retraerse a materia
no viva cuando sus condiciones no son realizadas. Pero la vida org�nica es definida
como procesos: no puede ser no din�mica, y esa es la raz�n por la cual cualquier
observaci�n de sus componentes caracter�sticos no basta para averiguar que haya
vida. �La vida no se puede definir mec�nicamente: podr� representar un mecanismo
en su construcci�n est�tica, pero nunca en su proceso evolutivo din�mico�, observaba
Jakob (1948).
Dicho
con brevedad, la sola presencia de componentes de vida no hace y no puede
demostrar la existencia de vida. Ni la formaci�n no biol�gica o inorg�nica y
los intercambios f�sicos de los componentes que en la Tierra operan como biomarcadores,
ni la acreci�n deposicional similar a la degradaci�n o fosilizaci�n de antiguos
organismos vivientes que en la Tierra ocurre en los combustibles f�siles, pueden
ser exclu�dos sin m�s. Por lo tanto, en base a razones epistemol�gicas, ning�n
sofisticado examen destinado a identificar mol�culas claves asociadas con la
vida puede brindar evidencia ninguna de vida genuina que sea, por principio,
algo m�s que tan s�lo entusiasmante, hasta sugerente: nunca podr�a ser
cient�ficamente concluyente.
Un sumario
de los experimentos de detecci�n de vida en la Misi�n Vikingo
Como
es sabido, los m�dulos de descenso de la Misi�n Vikingo llevaron varios
experimentos dise�ados no s�lo para descubrir materiales org�nicos, sino
tambi�n la existencia de organismos metabolizantes en la superficie marciana.
Un procedimiento confiable de detecci�n apuntado expresamente a este objetivo
no afronta ninguna objeci�n epistemol�gica, o de principio, en cuanto a su
capacidad de atribuir a organismos vivos sus resultados eventuales.
Mientras
un experimento no descubri�, en el suelo, compuesto org�nico ninguno, otro
grupo de nueve ensayos realizados, llamados en conjunto el experimento de
Liberaci�n Marcada (Labeled Release,
LR), encontr� resultados positivos. El LR fue dise�ado para dejar caer una
soluci�n nutritiva marcada con 14C en una peque�a muestra tomada del
suelo de Marte, monitoreando luego la evoluci�n de gas radiactivo: a saber, si
los microorganismos estaban metabolizando cualquiera de las sustancias nutritivas
que se les hab�a dado. Cuando el experimento fue conducido en ambos m�dulos de
descenso Vikingo, esto dio resultados positivos casi inmediatamente.
Algunos
cient�ficos llegaron a la conclusi�n de que los resultados positivos eran
atribuibles a �xidos en el suelo, que generaban una reacci�n qu�mica abi�tica
(es decir, no biol�gica) cuando la soluci�n nutritiva fue mezclada con los
�xidos. Sin embargo, el dise�ador del LR e investigador principal, Gilberto V.
Levin, estaba y sigui� estando convencido de que su experimento descubri� vida.
En los t�rminos de la susodicha definici�n adoptada por mi tradici�n neurobiol�gica,
esto significa que Levin afirma que la reacci�n qu�mica con la soluci�n
nutritiva es una parte del conjunto de
procesos eficientemente causales dependientes de su propia organizaci�n que, en
ciertos escenarios o ambientes, le permiten a esa organizaci�n continuar
existiendo. Levin tambi�n dice que el otro experimento, aquel que no
encontr� materiales org�nicos en el suelo, ni buscaba la detecci�n de vida activa
ni era bastante sensible para revelar materiales org�nicos en peque�as
cantidades, aserci�n que no fue controvertida y hoy por hoy refleja el acuerdo
general corriente.
Levin
2006 retrata as� la situaci�n reciente: "El 30 de julio de 2006 se cumpli�
el trig�simo aniversario del primer experimento de liberaci�n marcada para la detecci�n
de vida en Marte, llevado a cabo por la misi�n Viking. La potente respuesta
obtenida, junto a resultados concordantes de otros ocho ensayos adicionales de
liberaci�n marcada sobre suelo marciano, estableci� la presencia de un agente
activo que resultaba inhibido por el calentamiento. Los datos satisficieron los
criterios previos a la misi�n para la detecci�n de microorganismos vivientes.
Sin embargo, la comunidad cient�fica reaccion� con reserva, concluyendo en
general que la actividad constatada en el suelo tuvo causa qu�mica o f�sica. En
estas tres d�cadas la investigaci�n de Marte progres� mucho. � Empero, esa
vasta colecta de nueva informaci�n astrobiol�gica est� a�n por integrarse a una
evaluaci�n cient�fica objetiva de los resultados de la liberaci�n marcada en
Marte y de las posibilidades de vida all�. En realidad, en parte debido a
impropias interpretaciones de los nuevos hallazgos, algunos mitos han venido a
incrustarse en la literatura cient�fica sobre Marte. Tomando a esos mitos como
ingredientes clave, se ha desarrollado un equ�voco 'modelo tradicional' del
potencial de vida marciano. Lo acept� buena parte de la comunidad astrobiol�gica
y, por v�a de su endoso, tambi�n el ancho mundo. Este art�culo procura
presentar juntos los hechos sostenibles, buscando una revisi�n de ese consenso
actual concerniente a la vida en Marte. Recomienda asimismo acciones para facilitar
el cambio de modelo."
En
tales circunstancias, a fortiori
reexaminadas con los datos tra�dos al foco del escrutinio en los cuatro meses
que van desde mediados de octubre de 2006 a mediados de febrero de 2007, un
bi�logo sistem�tico podr�a preguntarse c�mo tratar correctamente las nueve
respuestas positivas, en dos grupos separados por aproximadamente 6000
kil�metros sobre la superficie de Marte, obtenidas por experimentos para la
detecci�n de vida realizados por ambos m�dulos de descenso de la Misi�n
Vikingo. Pienso que las dificultades conceptuales encontradas para manejar las
observaciones en t�rminos taxon�micos podr�an haber jugado un papel en la atribuci�n,
que el cient�fico responsable del experimento se�ala como injustificada, de
aquellos resultados a reacciones abi�ticas, esto es, s�lo f�sicas o qu�micas.
En los p�rrafos siguientes
relatar� datos conocidos y las observaciones mismas, como recientemente fueran
resumidas por el mencionado cient�fico y reproduci�ndolas aqu� con la debida
autorizaci�n, en beneficio de lectores transdisciplinarios que vinieren de
otros campos; los lectores bien informados al respecto podr�an m�s bien saltear
esta secci�n. La demostraci�n de metabolismo activo era el objetivo y con ello
la estructura conceptual del experimento de detecci�n de vida de liberaci�n
marcada. Un diagrama simple de este experimento se muestra en la Figura 1.
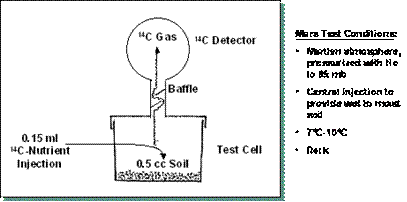
FIGURA 1.�
Esquema del experimento Viking de liberaci�n marcada en atm�sfera
marciana, presurizada con helio hasta 85 mb, en la oscuridad y 7 �C a 10 �C. Test cell, c�mara de prueba; soil, muestra recogida de suelo. Tomado
de Levin 2006a, b.
El
experimentador (Levin 2006a, b) expresa: "Todos los nutrientes, o substratos,
fueron compuestos moleculares simples, como los empleados en experimentos del
tipo Miller-Urey. Se los estima de temprana formaci�n en la Tierra primitiva y,
por eso, verosimilmente incorporados en las primigenias formas de vida, y probablemente
retenidos a lo largo de su proceso evolutivo. Cada nutriente candidato fue
uniformemente marcado con 14C. Esos nutrientes tienen is�meros
�pticos, por lo que fueron inclu�dos en forma de mezclas rac�micas a fin de que
cualquiera de sus dos estereois�meros estuviera disponible para incorporarse a
la potencial vida marciana. Los nutrientes fueron empleados en concentraciones
m�nimas en soluci�n de agua pura, para impedir alguna posible toxicidad como
sucede, a veces, cuando a los microorganismos se les proporciona liberalmente
materias org�nicas o inorg�nicas. La Tabla 1 presenta los nutrientes de liberaci�n
marcada, sus concentraciones y sus actividades."
TABLE 1.� Nutrientes de liberaci�n marcada
|
|
Estructura y
posici�n de |
|
|
Actividad
espec�fica |
|
14C-glicina |
NH3�*CH2�*COOH |
2.5 � 10-4M |
4 |
16 |
● Total = 34 μCi, que proporciona 6.8 � 107
dpm ml-1
"Miles
de ensayos fueron realizados con especies microbianas, cubriendo todo tipo
disponible: cultivo puro, cultivos mezclados y suelos; y numerosos ensayos de
campo con suelos fueron conducidos en un amplio rango de ambientes, durante los
veinte a�os de desarrollo del experimento de liberaci�n marcada. � Nunca se obtuvieron
falsos positivos a partir de muestras esterilizadas. La certidumbre de que la
respuesta proviene de organismos vivientes, la sensitividad (Levin y Straat
1979) (a apenas ~30 individuos o c�lulas/g), y la rapidez de la respuesta brindaron
alto nivel de confianza en el experimento."
En
cuanto a la historia del experimento
de liberaci�n marcada, su autor relata que "Una propuesta no solicitada
para desarrollar el experimento de liberaci�n marcada (originalmente,
'Gulliver') fue sometida a la NASA en 1958. Tras puntillosa revista, la
propuesta obtuvo fondos en 1959. El experimento de inmediato se revel� prometedor,
lo que fue detallado en informes trimestrales y anuales sometidos a la NASA.
A� fin de lograr su continuidad, una
nueva propuesta deb�a someterse anualmente a la NASA para su revisi�n. Hubo as�
constante interacci�n con la NASA durante todo el desarrollo. El proyecto Viking
fue formado en 1969 y entonces la NASA convoc� a una competici�n para experimentos
destinados a la detecci�n de vida. Muchas propuestas fueron presentadas, entre
ellas la de la liberaci�n marcada, que por otra vez atraves� el proceso de
evaluaci�n. El experimento de liberaci�n marcada fue seleccionado por los
cuatro comit�s de revisi�n establecidos por la NASA, cuyos miembros inclu�an personal
de la misma NASA, la National Science Foundation (NSF), los Institutos
Nacionales de la Salud (NIH) y las universidades. Todos ellos aceptaron los
criterios para detectar vida de la propuesta de liberaci�n marcada, a saber:
evoluci�n de gas marcado con 14C, seguida de un control tratado con calor
que produjese poco o nada de gas. Revisiones intensivas de la liberaci�n marcada,
programadas y de sorpresa, fueron llevadas frecuentemente a cabo por los comit�s
de la NASA y del Proyecto Viking as� como por 'tiger teams' (equipos especiales
para inspecci�n y evaluaci�n independiente) durante los �ltimos diez a�os de
desarrollo, todo lo cual aument� m�s aun el alto nivel de confianza que ten�an
sus numerosos revisores en el experimento de LR."
El
experimentador resume la ejecuci�n
as�. "Tras
posarse de modo impecable, Viking 1 llev� a cabo el primer experimento de liberaci�n
marcada el 30 de julio de 1976. El suelo que se somet�a a ensayo hab�a sido recogido
de la superficie por el brazo muestreador hasta una profundidad de unos cuatro
cent�metros, ubicado en la caja de distribuci�n y dispensado para la liberaci�n
marcada. De inmediato tras la inyecci�n de nutriente empez� un despliegue de
gas marcado con 14C. Despu�s de unos tres d�as de acumularse, el
incremento de volumen de dicho gas se aproxim� a una meseta, o plateau, aunque continu� mostrando un
aumento muy leve. Al final del ensayo del Ciclo 1 de ocho d�as marcianos o
soles, una segunda inyecci�n de nutriente fue efectuada. Se verific� una neta
disminuci�n del gas en el headspace o
espacio cabecera hasta que alrededor del 20% de dicho gas fue nuevamente
absorbido por la muestra, tras de lo cual una lenta re-evoluci�n del gas a lo
largo del Ciclo 2 de ocho soles restaur� la amplitud completa del Ciclo 1. El
protocolo requer�a control en caso de respuesta positiva. Acordemente, una
muestra duplicada de suelo fue insertada en una nueva celda, calentada por tres
horas a 160 oC para esterilizarla (el procedimiento establecido de
control para todos los experimentos Viking en biolog�a), se la dej� enfriar y
finalmente fue ensayada. La muestra duplicada virtualmente no produjo respuesta,
de modo que complet� los criterios pre-misi�n para la detecci�n de vida
microbiana. � M�s aun, aislada la muestra en la oscuridad
y mantenida en la caja de distribuci�n a ~10 oC, los ensayos de
liberaci�n marcada mostraron que en un per�odo de dos o tres meses el suelo
perd�a su actividad. Pero se obtuvieron respuestas positivas de muestras de suelo
que, antes de la inyecci�n de nutriente, hab�an sido almacenadas en esas mismas
condiciones durante varios d�as. Todos los resultados de liberaci�n marcada del
Viking Lander 1 (VL1), como muestra la Figura [2], sustentan la presencia de
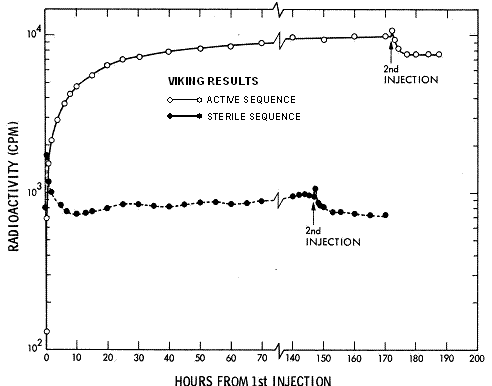
microorganismos vivientes o son consistentes con ella."
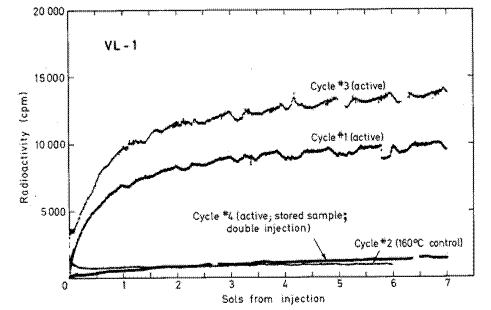
Figura 2. Todos los ciclos del VL 1. En la abscisa, soles desde la inyecci�n. La leyenda reza: Comparaci�n de la radioactividad desarrollada tras la primera inyecci�n de nutrientes radioactivos a cada ciclo de an�lisis del VL1. Una nueva muestra fue empleada para las secuencias activas de los ciclos 1 y 3 mientras la muestra utilizada para el ciclo activo 4 fue almacenada durante aproximadamente 141 soles a 10-26 �C antes de usarla. Para el ciclo 2, una porci�n almacenada de la misma muestra empleada para el ciclo 1 fue calentada durante tres horas a 160 �C antes de la inyecci�n de nutrientes. Todos los datos fueron ajustados seg�n los conteos de fondo observados antes de la inyecci�n. De Levin 2006a, b.
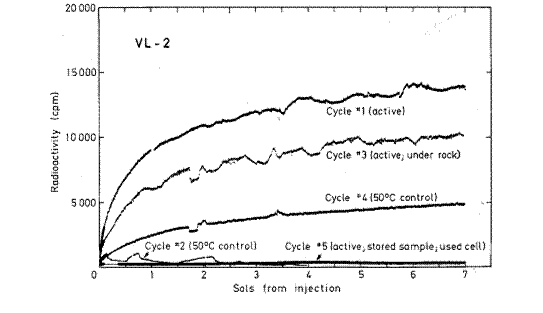
Figura 3. Todos los ciclos del VL 2. La leyenda reza: Comparaci�n de la radioactividad desarrollada tras la primera inyecci�n de nutrientes radiactivos a cada ciclo de an�lisis de VL 2. Una muestra nueva, fresca, fue empleada en cada ciclo, excepto el ciclo 5 que utiliz� una muestra almacenada antes de la inyecci�n durante aproximademente 84 soles a 7 �C. La muestra usada en el ciclo 3 fue obtenida de abajo de una roca. Los ciclos 1, 3 y 5 fueron secuencias activas, en tanto que los ciclos 2 y 4 fueron secuencias de control en los cuales las muestras furon calentadas por tres horas a ~ 61,5 �C y ~46 �C, respectivamente, antes de la inyecci�n de nutrientes. Los vol�menes de las muestras fueron de 0,5 cm3 excepto en el ciclo 5, que conten�a 2,2 cm3. Todos los datos fueron ajustados para los conteos de fondo observados antes de la inyecci�n. De Levin 2006a, b.
"A seis mil kil�metros de
distancia se posaba la sonda Viking 2. Sus resultados en materia de liberaci�n
marcada fueron muy similares a los de la VL1. En base al conocimiento adquirido
con los resultados de liberaci�n marcada de la sonda Viking 1, se ejecutaron controles
aun m�s definitorios para discriminar suplementariamente la naturaleza del agente activo. Estos controles incluyeron mover una
roca para permitir tomar una muestra de suelo que no hubiera estado expuesta a
la radiaci�n UV durante intervalos geol�gicos. Su activa respuesta refut� una
hip�tesis inicialmente prevalente, que sosten�a que la respuesta de liberaci�n
marcada era causada por la activaci�n UV del suelo. Aun otro ensayo demostr� que un calentamiento incluso modesto del suelo deprim�a
significantemente su respuesta. El agente activo en el suelo,
inicialmente respondiente a 10 oC, fue inhibido mucho o inactivado
por calentamiento a 46 oC o a 51 oC, tal como lo es una
variedad de microorganismos terrestres cuando se los sujeta a similar diferenciaci�n
termal (por ejemplo, E. coli v. otros
coliformes). Tal como en VL1, el almacenamiento del suelo durante meses en la
caja de distribuci�n inactiv� al agente. Todos los resultados de la liberaci�n
marcada de VL2 se muestran en la Figura [3].
Como en VL1, todos los resultados sostienen la presencia de microorganismos vivientes
o son consistentes con esa presencia."
En
este punto Levin, sin usar los modelos que concluyen que dentro de varios
metros a contar desde la superficie las esporas carentes de mecanismos para reparar
los da�os producidos por la radiaci�n penetrante podr�an empero sobrevivir durante
4,5 x 105 a�os (c�lculo publicado el 30 de enero de 2007 por Dartnell et al. 2007; ver tambi�n m�s abajo) comenta los
�bices puestos contra la aceptaci�n de los datos LR como prueba de vida, y los
problemas con cada uno, como sigue:
"a. Falla en
detectar materia org�nica. El instrumento de an�lisis org�nico de la misi�n Viking, un recortado
cromat�grafo de gases y espectr�metro de masa (GCMS, acr�nimo de gas chromatograph-mass spectrometer)
dise�ado para identificar el material org�nico que muchos presum�an habr�a de
hallarse presente en Marte, no encontr� mol�culas org�nicas (Biemann et al., 1977).� Con base en este resultado, el firme consenso
de la comunidad de ciencias del espacio fue que las respuestas positivas de la
liberaci�n marcada fueron de origen no biol�gico. Empero, el Experimentador
GCMS excluy� su instrumento como detector de vida, manifestando que para
obtener resultados hubiera sido requerida la cantidad de materia org�nica de 1000 millones de c�lulas
bacterianas (Biemann 1976). Ulteriormente, se inform� (Levin, Kuznetz and Lafleur, 2000) que varios problemas con el tipo de
instrumento GCMS del vuelo disiparon adicionalmente su sensitividad. Tras anunciarse
la detecci�n de materia org�nica en el meteorito marciano ALH4001, un funcionario
de la NASA explic� que el GCMS de la Viking no hab�a sido suficientemente sensitivo
para detectar el nivel de substancia org�nica encontrado por el instrumento
GCMS de escala completa con el cual se hab�a analizado el meteorite marciano.
Ha sido tambi�n mostrado (Benner 2000) que la temperatura
aplicada en el GCMS de la misi�n Viking no alcanzaba la magnitud necesaria para
vaporizar algunas mol�culas org�nicas estables a ese calor en c�lulas vivientes,
lo cual, se ha afirmado, podr�a explicar el resultado negativo del GCMS de la
misi�n Viking en detectar materia org�nica. A los instrumentos GCMS de las
sondas planetarias subsecuentemente dise�adas se les han incorporado correcciones,
dise�adas para reparar ese problema y as� proveerles una sensitividad muy incrementada.
Es interesante notar que la misi�n Viking, por s� misma, produjo evidencia de
que constantemente se est� formando materia org�nica en Marte; y de que la
misma no es destru�da por alg�n oxidante en�rgico. En efecto, los Experimentadores
de la liberaci�n pirol�tica (PR, Pyrolytic
Release) informaron (Horowitz, Hobby, and Hubbard,
1977): 'Los datos
muestran que una fijaci�n de carbono atmosf�rico tiene lugar en el material de
superficie de Marte bajo condiciones que se approximan a las marcianas.' En el
experimento, suelo marciano fue expuesto a atm�sfera marciana simulada conteniendo
CO2 y CO marcados. Tras 120 horas, cualquier gas carb�nico no fijado
fue apartado por medio de calor. Luego, tras calentar dicho suelo a temperatura
de pir�lisis, se vaporiz� en el espacio de cabecera cualquier carbono que
hubiera sido fijado. Cantidades estadisticamente significativas de gas
carb�nico marcado fueron desarroll�ndose a partir del material de suelo
marciano, proveyendo evidencia de que fijaci�n hab�a ocurrido (pero en cantidad
insuficiente para sostener la pretensi�n de que se trata de biolog�a). Esta
formaci�n de materia org�nica y su persistencia todo a lo largo del experimento
son evidencia contra la presencia del oxidante(s) o cualquier otra caracter�stica
del suelo que hubiese de destruir toda traza de materia org�nica. El
Experimentador de la liberaci�n pirol�tica hab�a informado (Hubbard et al., 1973 que 'Nuestros hallazgos
sugieren que el UV que al presente alcanza la superficie marciana puede estar
produciendo materia org�nica . . . seg�n lo encontrado, la cantidad de producto
podr�a ser considerable en intervalos geol�gicos.' "
"b. Oxidante en�rgico. Cuando, como arriba se indic�, el muestreo del
suelo debajo de una roca en Marte demostr� que la radiaci�n UV no es
responsable de las aparentes ausencias de vida y de materia org�nica, en su
lugar se propuso la presencia de per�xido de hidr�geno u otro(s) oxidante(s)
en�rgico(s) en el suelo. Tal hip�tesis fue formulada pese a los hallazgos del
experimento de propiedades magn�ticas de la misi�n (Figura 7 [no reproducida
aqu�]),
de que el
material de la superficie de Marte contiene un abundante componente magn�tico,
lo que es evidencia contra cualquier condici�n altamente oxidada (Hargraves et al.
1977). Los autores
del trabajo sobre el experimento Viking de propiedades magn�ticas llegaron a la
siguiente conclusi�n: 'Las posibilidades en cuanto a la naturaleza de las part�culas� magn�ticas detectadas en Marte se resumen
aqu�. Algunas o todas podr�an ser (1) granos de mineral sin oxidar altamente
magn�tico (Fe met�lico, magnetita, pirrotita) formando el n�cleo, bajo un
revestimiento rojizo de limionita o hematita'; y siguieron a�adiendo otras
varias posibilidades menores, ninguna de las cuales podr�a tornar altamente
oxidante el material de superficie. La evidencia contra un oxidante provista
por el PR, que antes acab�bamos de revisar, tambi�n fue despreciada por los
teorizadores pro-oxidante. Despu�s de la misi�n Viking, dos observaciones infrarrojas
(IR) con base en la Tierra, efectuadas por el orbitador ESA (Kerr 2004), y aun m�s recientemente datos del explorador rob�tico Opportunity
(Figura 8 [no
reproducida aqu�]), han
mostrado que el hierro en la superficie de Marte no est� oxidado por completo
(forma f�rrica), sino que ocurre mayormente en forma ferrosa. Por ende es arduo
defender la existencia de alg�n ubicuo oxidante en�rgico que destruya toda
materia org�nica en la superficie de Marte, o la sola presencia de dicho oxidante
en�rgico en ambos sitios de descenso de las sondas Viking, obligando a explicar
no biol�gicamente los resultados positivos en los ensayos de liberaci�n
marcada."

Figura 3. Comparaci�n de las respuestas activas terrestres y marcianas a la LR. De Levin 2006a, b.
"c. 'Demasiado, y demasiado pronto.' Se ha objetado que las respuestas positivas
a la liberaci�n marcada, as� como la cin�tica de la reacci�n, corresponden a
una reacci�n de primer orden, sin la demora o fases exponenciales que se observan
en las cl�sicas curvas de crecimiento microbiano � todo lo cual hablar�a de una
simple reacci�n qu�mica. Empero, la Figura [3, arriba] muestra experimentos terrestres
de liberaci�n marcada en una variedad de suelos, que produjeron tasas de
respuesta con la cin�tica y el rango de amplitudes de las liberaciones marcadas
en Marte. d. Segunda Inyecci�n. Las segundas inyecciones de
nutrientes no produjeron nuevo desarrollo de gas. Al contrario, prestamente
redujeron en alrededor del 20% la cantidad de gas acumulado desde la primera
inyecci�n. Si bien la responsividad a las segundas inyecciones no form� parte
de los criterios para la detecci�n de vida por medio de liberaci�n marcada, la
ausencia de un nuevo surgimiento de gas al inyectar medio fresco fue
posteriormente citada como evidencia contra la biolog�a. Empero, un ensayo de
suelo consolidado provisto por la NASA (Antarctic
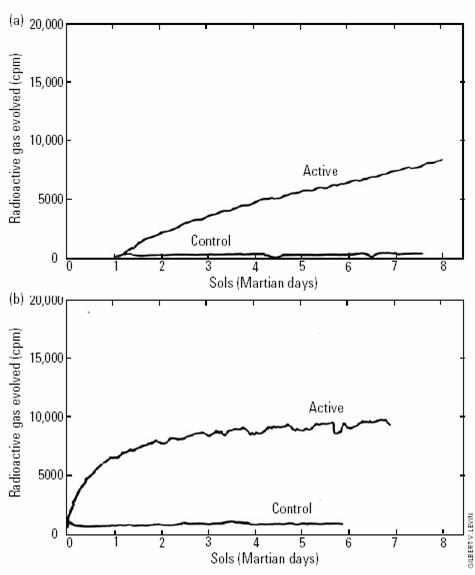
soil No. 664), conteniendo menos de 10 c�lulas viables/g (Quam 1971), mostr� este tipo de respuesta�
a la� 2da inyecci�n,
como se observa en las Figures 10a y 10b [aqu� Fig. 4a y
4b ]. � As�, pues,
el fracaso de la 2da inyecci�n en elicitar una respuesta puede atribuirse
a que los organismos en la muestra activa hubiesen muerto en alg�n momento tras
la 1ra inyecci�n, durante la �ltima etapa del Ciclo 1. El efecto de
la 2da inyecci�n fue humedecer el suelo, causando que el mismo absorba
gas del espacio de cabecera. El gradual resurgir, con el tiempo, de gas en
dicho espacio parece haber sucedido a medida que el sistema se puso en
equilibrio."
La secci�n
siguiente de Levin 2006a, b ha sido abreviada aqu�, porque la disponibilidad de
cantidades importantes de agua helada subsuperficial ya dej� de ser contenciosa
y ello vari� la plausibilidad de los escenarios de Levin: "e. 'No hay agua l�quida, no hay vida.' Esta alegaci�n es el argumento primario interpuesto
hoy por hoy por quien no acepte el descubrimiento de vida por la liberaci�n
marcada de las Viking. Sin embargo, la misma misi�n Viking brind� firme evidencia (Moore et al. 1977) de la presencia de agua l�quida cuando el aumento
de temperatura de su pie, respondiendo al levantarse del sol, se detuvo a 273 oK.
Se ve nieve o helada en las im�genes de la misi�n Viking del sitio de descenso
(Figura 11 [Viking Lander Imagen 21I093, no reproducida
aqu�]). � Explicando la pegajosidad
del suelo, cient�ficos del MER han dicho que 'puede contener peque�os gl�bulos
de agua l�quida', o 'puede contener fangos� (brine)'."
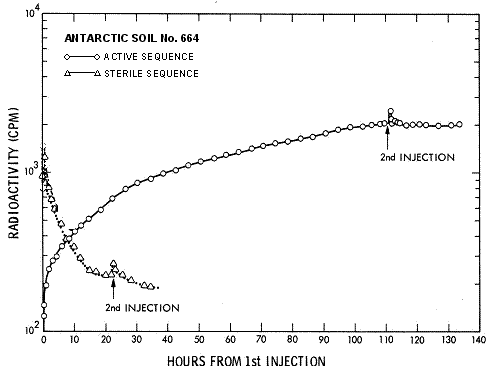
FIGURA
4a.� Efecto de la 2da
inyecci�n en suelo ant�rtico. De Levin 2006a, b.
FIGURA
4b.� Efecto de la 2da
inyecci�n en suelo marciano. De Levin
2006a, b.
"Otras im�genes de Marte, � muestran
la actividad presente, aunque intermitente, de regueros o arroyuelos. � en los hielos del Polo Sur terrestre � y en
el permafrost del �rtico � aun en esos congelados sitios, existe agua l�quida.
Pel�culas muy delgadas de agua l�quida existen en los intersticios del hielo y
minerales, y bastan para sostener una ecolog�a de especies altamente
diferenciadas. Barniz del desierto. En 1979 se llam� la atenci�n
del autor (Henry Moore
a Gilberto Levin, comunicaci�n personal) sobre la posible presencia del denominado barniz del desierto en algunas de las
rocas marcianas. �� [noticias recientes]� � informan acerca de un reencendido inter�s en el
barniz del desierto como evidencia de la vida en Marte."
FIGURA
5.� Curvas de
crecimiento microbiano tras aplicar la soluci�n nutriente marcada a suelo del desierto
californiano y su control tratado por calor (arriba) y el resultado paralelo
del m�dulo de descenso Vikingo 1. En ambos casos las muestras de control
fueron calentada a 160 �C por tres horas. Gentileza G.
V. Levin.
"Ritmo circadiano. El re-examen de la cin�tica de la
respuesta a la liberaci�n marcada en Marte indic� otra posible componente
biol�gica. Se ha propuesto (Levin et al, 2004; Van Dongen et al., 2005) �que
la cin�tica de evoluci�n del gas marcado en los experimentos Viking de
liberaci�n marcada podr�a atribuirse a un ritmo circadiano, un fen�meno
biol�gico casi universal en todo organismo vivo. Mientras que se detectaron indicaciones
de ritmo circadiano en los datos Viking de liberaci�n marcada, en los dos
art�culos citados las mismas no alcanzaron el nivel de firme significaci�n
estad�stica. Empero otro art�culo (Bianciardi
2004), utilizando una aproximaci�n
no linear, concluy� as�: 'Nuestros resultados apoyan con firmeza la hip�tesis
de un origen biol�gico del gas colectado por el experimento de liberaci�n
marcada a partir de suelo marciano.' Estudios adicionales al presente en progreso
se encaminan a verificar la significaci�n estad�stica de esa conclusi�n. Indicadores atmosf�ricos. Sum�ndose a
la creciente marea de hechos que sostienen la detecci�n de vida por los experimentos
Viking de liberaci�n marcada, se encuentran los recientes hallazgos, en la atm�sfera
marciana, de metano, formaldeh�do y posiblemente amon�aco � Estos gases con
frecuencia se implican en el metabolismo microbiano y son, en consecuencia, posibles
indicadores de vida. El metano, gas l�bil ante el UV y de breve vida media,
ocurre en cantidades estimadas no adecuadas para su tasa de reemplazo, ya que
el catastro termal de todo el planeta no ha indicado ninguna actividad
volc�nica, potencial fuente no-biol�gica de metano. En la atm�sfera de la
Tierra, el metano se sostiene primariamente por el metabolismo biol�gico. A
m�s, el metano detectado en Marte estaba asociado con vapor de agua en la
atm�sfera, lo que es consistente con la posible existencia de vida, si no indicativo
de la misma."
Consideraciones
de conjunto sobre los experimentos Vikingo
El 22
de marzo de 2007, un seminario a�n impublicado sobre "La argumentaci�n
acerca de la vida en Marte" en la Washington State University School of Earth and
Environmental Sciences reuni�
entre otros a Levin y Schulze-Makuch. [Se prevee continuar con un nuevo seminario
en la Carnegie Institution de Washington,
el pr�ximo 14 de mayo. N. d. T.] Estos nuevos an�lisis de los resultados de la
Misi�n Vikingo concluyeron afirmativamente sobre un Marte biosfereado, desplaz�ndose
consiguientemente el foco del an�lisis a evaluar la plausibilidad de la
asunci�n de Houtkooper y Schulze-Makuch. Se trata de la hip�tesis de vida
marciana en base al solvente H2O2-H2O, a saber
la hip�tesis de que uno de los solventes internos de organismos vivientes en
Marte podr�a ser H2O2-H2O m�s bien que H2O.
Las proporciones que, a la fecha de redactar el presente trabajo, est�n siendo
consideradas como las m�s prometedoras para examinar, se centran en un 26 % H2O2
y un 74 % H2O, con el H2O2 en los l�mites del
17 % al 35 %. Los citados autores (Houtkooper y Schulze-Makuch 2007) argumentan
que las condiciones ambientales corrientes cerca de la superficie de Marte no
son incompatibles con la vida, asumen la existencia probable de organismos
vivos, y as� proponen "una reinterpretaci�n de los resultados de la Misi�n
Vikingo, basada en suponer que los microorganismos sobre Marte produzcan el per�xido
de hidr�geno y generen un solvente intracellular de H2O2-H2O
para los procesos bioqu�micos seleccionados por el particular ambiente marciano
y adaptados al mismo."
La
hip�tesis, de que los organismos marcianos utilizar�an una mezcla H2O2-H2O
como l�quido intracellular, da cuenta de los resultados del experimento de
liberaci�n marcada asumiendo que los organismos vivos ya hab�an fallecido en el
momento de la segunda inyecci�n nutritiva, debi�ndose la absorci�n del CO2
antes desarrollado (un metabolito) a la alcalinidad
del suelo humedecido (J. Houtkooper, D. Schulze-Makuch, comunic. personal a
quien escribe).
Cierto
n�mero de asuntos relacionados con esta asunci�n a�n no han sido totalmente
resueltos ni calculados, como las coordinaciones hidrof�lico-hidrof�bicas en la
formaci�n de las membranas (que es un proceso continuo, de equilibrio), o el papel
(sugerido por quien escribe) en compartimentar vol�menes restringidos de cualquier
mezcla H2O2-H2O, que una vez supuesta tal
hip�tesis pueden jugar capas de agua estructurada (probablemente tambi�n v�trea
m�s que disruptivamente cristalina a las temperaturas relevantes debajo de 0
�C) adsorbida sobre an�logos del citoesqueleto y organelas. Sin embargo, aqu� lo
esencial de esa contribuci�n de Houtkooper y Schulze-Makuch � y lo que refleja
los cambios recientes � es su asunci�n de vida marciana.
Respecto
a la alcalinidad hipotetizada para el suelo marciano, otro hecho nuevo de relevancia
es el se�alamiento de que, si el color rojizo del suelo marciano ya no pudiera
atribuirse del todo a que la superficie est� mayormente oxidada, tal hecho del
color rojizo podr�a en cambio atribuirse a la reflexi�n de una profusa absorci�n
de otras bandas del espectro por microorganismos fotosint�ticos.
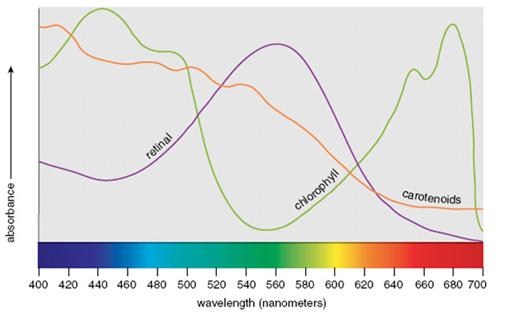
FIGURA
6.� Las curvas en
relaci�n especular han sugerido a algunos que la clorofila evolucion� para explotar
bandas espectrales no utilizadas por el retinal, que podr�a haber sido general en
las bacterias previamente dominantes. Los lagos p�rpura de Australia toman su color
de halobacterias que emplean retinal para capturar la luz solar.
Tal
se�alamiento fue efectuado en el congreso de la European Geosciences Union en Viena por el ge�logo ambiental y paleoclimat�logo
holand�s Roland Paepe (2007), quien en la oportunidad resalt� que una "observaci�n desde cerca de las
huellas de impacto en el sitio del m�dulo as� como las reciente detecci�n de
arcillas de filosilicatos en Marte, puede llevar a indicaciones firmes de
procesos pedogen�ticos ... Ello abre nuevas posibilidades para el estudio del
desarrollo de suelos similares a los procesos de suelo terrestres � ahora que
los filosilicatos han sido detectados en Marte, el rol del agua en el proceso
clim�tico de transformaci�n de suelos arcillosos ha sido indudablemente probado.
Esto puede implicar adem�s que no s�lo el agua y la transformaci�n clim�tica o
pedog�nesis se extienden sobre Marte, sino tambi�n una cubierta de [microorganismos].
La excesivamente general connotaci�n de 'suelo' debe pues ser reformada dando
lugar al genuino concepto de� suelo, que
envuelve agua y vegetaci�n en su desarrollo. Por ello la relaci�n entre desarrollos
de suelo y vida vegetal o bacteriana en la superficie de Marte abre nuevas y amplias
posibilidades para las investigaciones en astrobiolog�a. Los suelos en el espacio
y su contenido cianobacterial asociado han de tornarse foco para investigar
evidencias de real desarrollo de suelos en el sistema solar, fuera de nuestro
planeta."
Uno de
los condicionantes de ese desarrollo es el flujo de radiaci�n a trav�s del
suelo. Como ya fue mencionado, Dartnell et al. 2007 consideran el flujo entrante tanto de
part�culas de rayos c�smicos gal�cticos (GCR) como de protones solares
en�rgicos (SEP) sobre una amplia gama de energ�as, sumando dosis de radiaci�n.
Como resultado, concluyen, "encontramos que en 2 m de profundidad, que es
el alcance del taladro de ExoMars, una poblaci�n de c�lulas radioresistentes
tendr�a que haberse reanimado dentro de cada 450.000 a�os para seguir siendo
todav�a viable." Como comenta Levin (comunic. personal a M. Szirko), esto
significa que "ellos reconocen que la radiaci�n incidente en la superficie
de Marte es realmente trivial para la supervivencia inmediata de mucha especies
de microorganismos [de tipo terrestre] � incluso sin necesidad de invocar a
Darwin para realzar su protecci�n o tolerancia a lo largo de los
milenios", y "Todo parece conferir perspectivas muy buenas para la
supervivencia (como seguramente se demostr� cuando la c�mara del Surveyor fue tra�da
de vuelta de la Luna despu�s de permanecer cuarenta meses en sus campos de
radiaci�n, mucho m�s rigurosos que los de Marte, encontr�ndose que �a�n
conten�a microorganismos viables!)"
En
cuanto a una aserci�n de Dartnell et al. 2007 afirmando que el agua como l�quido es inestable sobre
la superficie marciana, Levin advierte (�b.)
que a lo largo de sus seis a�os de operaci�n, ning�n m�dulo de descenso Vikingo
refiri� una presi�n atmosf�rica de superficie por debajo del punto triple del
agua (6.1 mb), y que casi todo el vapor de agua sobre Marte est� muy cerca del
suelo, suministrando diurnamente una atm�sfera saturada sobre la mayor parte
del planeta. En tales circunstancias, como la evaluaci�n por Dartnell et al. 2007
no consider� ning�n mecanismo
de reparaci�n fisiol�gica que funcione adaptativamente en el metabolismo
activo, su ejercicio independiente de modelismo puede ser le�do como
significando que, no ya en profundidad sino hasta en la misma superficie de
Marte, la suma entrante de GCR m�s SEP no plantea biol�gicamente constre�imiento
serio ninguno a organismos vivos correctamente adaptados no-inactivos, o
reanimados a diario o hasta por estaciones del a�o.
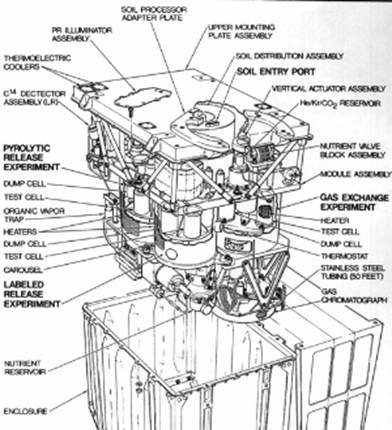
FIGURA
7.� Disposici�n de
los experimentos para detectar vida activa en los m�dulos de descenso de la
Misi�n Vikingo (NASA).
La acci�n
nomenclatural
Como lo
expresan Houtkooper y Schulze-Makuch (2007), "El argumento de que la misi�n
Vikingo descubri� reacciones qu�micas m�s bien que biolog�a, basado en el hecho
de que no hay organismos terrestres que puedan reproducir todos los resultados
de Vikingo, est� errado. De la misma manera lo est� el argumento de que la misi�n
Vikingo descubri� biolog�a conocida, porque en la Tierra no hay ninguna qu�mica
mineral o reactiva conocida an�loga que produzca todos los resultados de
Vikingo. Cualquier explicaci�n de los resultados de la misi�n Vikingo debe ser
intr�nsecamente vinculada al ambiente marciano � " Esta situaci�n, as�
como las circunstancias de Historia de las Ideas y de sociolog�a de la ciencia
mencionadas en la Introducci�n y sus secciones consiguientes, y las
reinterpretaciones significativas de los experimentos Vikingo en los seis meses
pasados bajo el nuevo conocimiento acumulado, tornan obsoleta la advertencia postulada
el seis de septiembre de 1976, por Klein et al. en su art�culo de Science 'The Viking Biological Investigation: Preliminary Results' � donde preven�an que "Interpretaciones
alternativas, qu�micas y biol�gicas, son posibles para estos datos
preliminares. Los experimentos est�n todav�a en proceso y sus resultados hasta
ahora no permiten una decisi�n en cuanto a la existencia de vida sobre el
planeta Marte" � y m�s bien requieren ampliar la biolog�a fuera de la
Tierra, describiendo y emplazando sistem�ticamente en la taxonom�a biol�gica al
agente activo marciano responsable de los resultados del experimento de Liberaci�n
Marcada de la Misi�n Vikingo de 1976.
Ello
as�, por la presente vengo formalmente a proporcionar la siguiente entrada
taxon�mica para el agente activo caracterizado por su comportamiento relatado
como responsable de los resultados del experimento de Liberaci�n Marcada de la
Misi�n Vikingo de 1976:
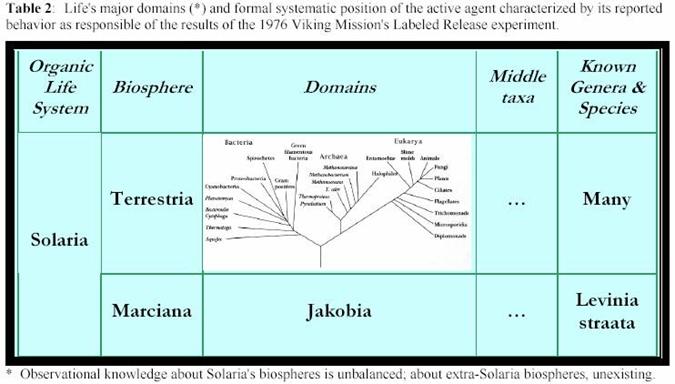
� Sistema
de vida org�nica, Solaria, novum vit� systema. Comprende todos los organismos vivientes
extintos y existentes en el Sistema Solar, organismos vivientes definidos en lo
que antecede� como la convergencia de
subprogramas que sostiene sistemas estoc�sticos multivariados, entendidos como
el conjunto de procesos eficientemente causales dependientes de la organizaci�n
de dichos sistemas que, en ciertos escenarios o ambientes, le permiten a esa
organizaci�n continuar existiendo.
� Bi�sfera,
Marciana, nova biosph�ra: todos los organismos vivientes extintos
y existentes cuyos linajes se desarrollaron sobre el planeta solar Marte. Su
tax�n paralelo es Terrestria, tambi�n novum
nomen: todos los organismos vivientes extintos y existentes cuyos linajes
se desarrollaron sobre el planeta solar Tierra, L. Terra.
� Reino,
Jakobia, novum regnus: todos los organismos vivientes
integrantes de Marciana cuya anatom�a y fisiolog�a est�n adaptadas para vivir
habitualmente en las gamas de radiaci�n ultravioleta y otras condiciones caracter�sticas
del suelo superior del Marte, incluso si fuesen tambi�n capaces de sobrevivir
en otro biotopos marcianos. El reino es as� denominado para honrar a Christofredo
Jakob (Christfried Jakob, 1866-1956), neurobi�logo que dedic� su vida al
esfuerzo de entender y conceptuar la vida.
� G�nero
y especie Gillevinia straata, novum genus et species nova. [Gillevinia, nuevo nombre de g�nero, substituye a Levinia; ver pp. 35-37]. Queda caracterizado por su relatado
comportamiento como responsable de los resultados del experimento de Liberaci�n
Marcada de la Misi�n Vikingo de 1976, en experimentos sobre las muestras de
material marciano superficial ("suelo") conducido dentro de los m�dulos
de descenso Vikingo y resumido en Klein et al.
1976 �por los rasgos siguientes: "En la
asimilaci�n de carbono o experimento de liberaci�n pirol�tica, 14CO2 y 14CO
fueron expuestos al suelo
en presencia de luz. Se encontr� una peque�a cantidad de gas convertida en
material org�nico. El tratamiento t�rmico de un duplicado de la muestra impidi�
tal conversi�n. En el experimento de intercambio de gas, el suelo primero fue
humectado (expuesto a vapor de agua) durante 6 soles y luego rehumedecido con
una soluci�n compleja acuosa de metabolitos. El gas encima del suelo fue monitoreado
por cromatograf�a gaseosa. Una cantidad sustancial de O2 fue detectada en el primer cromatograma tomado 2,8 horas despu�s de la
humidificaci�n. An�lisis subsecuentes revelaron que tambi�n hab�an ocurrido aumentos
significativos de CO2 y s�lo peque�os cambios en N2. En el experimento de liberaci�n marcada, el suelo fue humedecido con
una soluci�n que contiene varios compuestos org�nicos marcados con 14C. Un desarrollo sustancial de gas
radiactivo fue registrado, lo que no ocurri� con una muestra duplicada tratada
por calor." El g�nero y la especie son as� denominados para honrar a
Gilberto V. Levin, quien despu�s de trabajar dieciocho a�os en desarrollar y
llevar a vuelo el experimento de Liberaci�n Marcada de la misi�n Vikingo dedic�
otro esfuerzo de treinta a�os a adquirir los hechos cient�ficos y conclusiones
anal�ticas que apoyan la plausibilidad de la interpretaci�n biol�gica de este
comportamiento; y Patricia Ana Straat, co-experimentador del experimento de
Liberaci�n Marcada de la misi�n Vikingo, cuyos esfuerzos fueron indispensables
para el desarrollo del experimento y su vuelo.
� Holotipo:
como portador modelo del nombre nuevo, los restos de los espec�menes
originalmente ensayados se encuentran por ahora conservados naturalmente sobre
Marte (todav�a dentro de los dos m�dulos de descenso Vikingo, que han sido
debidamente localizados y puestos en imagen por la c�mara del Experimento Cient�fico
en Adquisici�n de Im�genes de Alta Resoluci�n del Orbitador de Reconocimiento
de Marte). La accesibilidad, como parece que habr� de ser t�pico para la
exobiolog�a, ser� en particular limitada mientras se desarrollan medios
apropiados y seguros para traer espec�menes biol�gicos a la Tierra. Aunque la
biomasa total de Marciana parezca considerablemente m�s peque�a que la de
Terrestria (dado que una biomasa a plena escala parece incompatible con la
condici�n pr�xima al equilibrio qu�mico y termodin�mico de la atm�sfera actual
de Marte), como inferencia bayesiana cabe suponer que ejemplares vivientes se
hallan distribuidos en otra partes sobre similares suelos marcianos. Por consiguiente
epitipos, esto es tipos adicionales y clarificantes, podr�an ser eventualmente
designados en lugar de los holotipos ensayados por la Misi�n Vikingo.
La
demarcaci�n de Gillevinia straata,
como es habitual en la sistem�tica de bacterias en Terrestria, no ha sido
guiada seg�n un concepto de especie basado teoricamente. Cohan 2002 se�ala que "una especie es un grupo
de organismos cuya divergencia es encapsulada por una fuerza de cohesi�n; la
divergencia entre especies diferentes es irreversible y diferentes especies son
ecologicamente distintas. En el caso de bacterias, estas propiedades
universales son sostenidas no por la denominada especie de la sistem�tica, sino
por ecotipos. Estos son poblaciones de organismos que ocupan el mismo nicho
ecol�gico, cuya divergencia es purgada recurrentemente por la selecci�n natural.
Tales ecotipos pueden ser descubiertos por medio de varias aproximaciones basadas
en secuencias universales. Estos m�todos moleculares sugieren que una t�pica
especie denominada contiene muchos ecotipos, cada uno con los atributos
universales de especie." Una situaci�n similar, con ecotipos que modelen
sus nichos y regulen los gradientes ambientales de modo de competir en
diversificaci�n, torn�ndose unidades de nivel superior para las presiones de
selecci�n natural, tambi�n puede ser de esperar para todo el reino Jakobia.-
��
Cr�ditos
El autor est�
profundamente agradecido a Jorge Telerman, Jefe de Gobierno de la Ciudad de
Buenos Aires, por haber proporcionado un apoyo crucial que permiti� la producci�n
del presente art�culo.
��
Referencias
Benner, S.
(2000), quoted in New Scientist article, �Buried Evidence,� March 24. (Apud
Levin 2006a, b).
Bianciardi,
G. (2004), �Nonlinear Analysis of the Viking Lander 2 Labeled Release Data,�
Proc. of the III European Workshop on Exo-Astrobiology on Mars: The search for
Life, Madrid, Spain, 18-20 November 2003 (ESA SP-545, March). (Apud
Levin 2006a, b).
Biemann, K.
(1976), Viking Science Press Conf. Sept. 4, JPL. (Apud Levin 2006a, b).
Biemann, K.
et al. (1977), �The Search for
Organic Substances and Inorganic Volatile Compounds in the Surface of Mars,� J.
Geophys. Res. 82, 28, 4641-4662. (Apud Levin 2006a, b).
Cameron,
R.E. and R.E. Benoit (1970), Ecology 51, 801. See also Antarctic Soil No. 715.
(Apud Levin 2006a, b).
Cohan, F.
M. (2002), "What are bacterial species?", Annual Review of
Microbiology 56: 457-487
doi:10.1146/annurev.micro.56.012302.160634
Crocco, M.
(2007), "A Palindrome: Conscious Living Creatures as Instruments of
Nature; Nature as an Instrument of Conscious Living Creatures", Chapter 12
in Helmut Wautischer, ed., Ontology of Consciousness, The MIT Press, Cambridge,
MA.
Dartnell,
L. R., L. Desorgher, J. M. Ward, and A. J. Coates (2007), "Modelling the
surface and subsurface Martian radiation environment: Implications for astrobiology",
Geophys. Res. Lett., 34, L02207, doi:10.1029/2006GL027494
Hargraves,
R.B., D.W. Collinson, R.E. Arvidson and C.R. Spitzer (1977), �The Viking Magnetic
Properties Experiment: Primary Mission Results,� J. Geophys. Res. 82, 4547 (Apud
Levin 2006a, b).
Houtkooper,
J. M. and D. Schulze-Makuch (2007), " A Possible Biogenic Origin for
Hydrogen Peroxide on Mars: The Viking Results Reinterpreted", in press at Int.
J of Astrobiology.
Horowitz,
N.H., G.L. Hobby and Jerry S. Hubbard (1977), �Viking on Mars: The Carbon Assimilation
Experiments,� J. Geo. Res. 82, 28, 4659-4662. (Apud Levin 2006a,
b).
Hubbard,
J.S., J.P. Hardy, G.E. Voecks and E.E. Golub (1973), �Photocatalytic Synthesis
of Organic Compounds from CO and Water: Involvement of Surfaces in the
Formation and Stabilization of Products�, J. Mol. Evol. 2, 149-166. (Apud
Levin 2006a, b).
Jakob, C.
(1948), "La definici�n cient�fica de la vida", Revista del Museo
Social Argentino XXXVI, issues 313-314, pp. 193-203, julio-agosto. Republished
with a Preliminary Notice by M. Szirko and a study by Ramon Carrillo
("�Qu� es la vida?"; all in Spanish) in Electroneurobiolog�a (2000) 7
(1), 1-43;
http://electroneubio.secyt.gov.ar/Christfried_Jakob_vida_definicion_cientifica.htm
Kerr, R.A.
(2004), �Life or Volcanic Belching on Mars?� Science 303, # 5666, 1953, 26
March (Apud Levin 2006a, b).
Klein, H.
P., N. H. Horowitz, G. V. Levin, V. I. Oyama, J. Lederberg, A. Rich, J. S.
Hubbard, G. L. Hobby, P. A. Straat, B. J. Berdahl, G. C. Carle, F. S. Brown,
and R. D. Johnson (1976), "The Viking Biological Investigation:
Preliminary Results", Science 194 (4260), 99-105. DOI: 10.1126/science.194.4260.99
Levin, G.
V. (2006a),"Modern Myths Concerning Life on Mars", Electroneurobiolog�a
14 (5), 3-25; http://electroneubio.secyt.gov.ar/Gilbert_V_Levin_Life_on_Mars_Modern_Myths.htm
Levin, G.
V. (2006b),"Mitos modernos acerca de la vida en Marte", Electroneurobiolog�a
14 (5), 27-52;
http://electroneubio.secyt.gov.ar/Gilbert_V_Levin_Vida_en_Marte_Mitos_Modernos.htm
Levin,
G.V.; P.A. Straat, H.P.A. Van Dongen, and J.D. Miller (2004), �Circadian
rhythms and evidence for life on Mars,� Instruments, Methods, and Missions for
Astrobiology, SPIE Proceedings 5555, 35, August (Apud Levin 2006a, b).
Levin, G.;
L. Kuznetz, and A. Lafleur (2000), �Approaches to resolving the question of
life on Mars,� Instruments, Methods, and Missions for Astrobiology, SPIE
Proceedings 4137, 48-62. (Apud Levin 2006a, b).
Levin, G.V.
and P.A. Straat (1979), �Completion of the Viking Labeled Release Experiment on
Mars�, J. Mol. Evol. 14, 167-183.
Moore, H.J.
et al. (1977), �Surface Materials of
the Viking Landing Sites,� J. Geophys. Res. 82:28, 4497-4523 (Apud Levin
2006a, b).
Navarro-Gonz�lez,
R; Navarro, K. F.; de la Rosa, J., I�iguez, E.; Molina, P.; Miranda, L. D. ;
Morales, P; Cienfuegos, E.; Coll, P.; Raulin, F., Amils, R. and McKay, C. P.
(2006), "The limitations on organic detection in Mars-like soils by
thermal volatilization�gas chromatography�MS and their implications for the
Viking results", PNAS 103 (44 ), 16089-16094. http://www.pnas.org/cgi/content/full/103/44/16089
Paepe, R.,
(2007), "The Red Soil on Mars as a proof for water and vegetation"
Geophysical Research Abstracts 9, 01794; SRef-ID:
1607-7962/gra/EGU2007-A-01794, http://www.cosis.net/abstracts/EGU2007/01794/EGU2007-J-01794.pdf?PHPSESSID=e�
Quam, L.O.,
ed. (1971), Research in the Antarctic, AAAS #93, Wash., DC. (Apud Levin
2006a, b).
Szirko, M.
(2007), "Comentario editorial: la cuesti�n epistemol�gica en la detecci�n
de vida en Marte", Electroneurobiolog�a 15 (1), 183-187;
http://electroneubio.secyt.gov.ar/Acerca_de_la_vida_en_Marte_Editorial.htm
Van Dongen,
H., J. Miller, P. Straat and G. Levin (2005), �A circadian biosignature in the
Labeled Release data from Mars?� Instruments, Methods, and Missions for
Astrobiology, SPIE Proceedings 5906, OC1-10, August (Apud Levin 2006a, b).
Copyright � April 2007 Electroneurobiolog�a. Este
texto es un art�culo de acceso p�blico; su copia exacta y redistribuci�n por
cualquier medio est�n permitidas bajo la condici�n de conservar esta noticia y
la referencia completa a su publicaci�n incluyendo la URL (ver primera p�gina).
/ This is an Open Access article: verbatim copying and redistribution of this
article are permitted in all media for any purpose, provided this notice is
preserved along with the article's full citation and URL (first page).

revista
Electroneurobiolog�a
ISSN: ONLINE 1850-1826 - PRINT 0328-0446
Correcci�n:
primera clasificaci�n biol�gica de un organismo marciano, g�nero Gillevinia (no Levinia)
Correction note: first biological classification of
a Martian organism, genus
Gillevinia
(not Levinia)
por / by
Mario
Crocco
Director, Centro de Investigaciones�
Neurobiol�gicas, Ministerio de Salud, Rep�blica Argentina, y Jefe de,
Laboratorio de Investigaciones�
Electroneurobiol�gicas, Hospital "Dr. J. T. Borda", Gobierno
de la Ciudad de Buenos Aires
Ave. Amancio Alcorta 1602,
1283 Buenos Aires, Argentina; tel./fax (54) 11 4306 7314, mcrocco[at]electroneubio.com.ar
Electroneurobiolog�a 2007; 15 (2), pp.
35-37; URL <http://electroneubio.secyt.gov.ar/index2.htm>
Copyright � April 2007 Electroneurobiolog�a. Este texto es un art�culo de acceso p�blico; su copia exacta y redistribuci�n
por cualquier medio est�n permitidas bajo la condici�n de conservar esta
noticia y la referencia completa a su publicaci�n incluyendo la URL (ver
arriba). / This is an Open Access article: verbatim copying and redistribution
of this article are permitted in all media for any purpose, provided this
notice is preserved along with the article's full citation and URL (above).
Publication date / fecha de publicaci�n:� 28 de abril, 2007
Se ha hallado que Levinia, el nombre brindado
el 20 de diciembre de 2006 al g�nero de espec�menes activos encontrados en la bi�sfera Marciana por el experimento
de liberaci�n marcada de la Misi�n Vikingo de 1976, cuyos detalles diagn�sticos
(characteres naturales) fueron provistos
por Klein et al.
1976, ya estaba en uso para g�neros de la bi�sfera Terrestria.
En efecto, en Trilobites se
emplea corrientemente (Jell y Adrain 2003) el nombre gen�rico Levinia, con el cual Rusconi (1950) nomencl� en Mendoza, Argentina,
un g�nero o subg�nero trilobite de familia indeterminada. Entre los "coccidios",
o protozoos par�sitos del suborden Eimeriorina en el phylum Apicomplexa, el
nombre gen�rico Levinia ha sido tambi�n
propuesto para las especies del g�nero Isospora
que se hospedan en especies parat�nicas (cf.
Dubey 1977). Entre las Enterobacteriaceae, Levinia es asimismo una variante de
escritura o bien sin�nimo de un tard�o g�nero heterot�pico (Levinea Young et al. 1971) a su vez sinonimial de
Citrobacter
Werkman y Gillen 1932 (cf. Farmer 1981, Brenner et al 1993). En
consecuencia, a fin de asegurar que los nombres de g�nero sean �nicos a
trav�s de todos los taxones biol�gicos mayores, el nombre de g�nero Levinia es reemplazado y el g�nero que el mismo caracterizaba
en Marciana es por
la presente nomenclado Gillevinia, novum genus. Por
ende, el agente activo caracterizado
por su oportunamente comunicado comportamiento como responsable de los
resultados del experimento de liberaci�n marcada en la Misi�n Vikingo de 1976 desde
ahora constituye formalmente, para la biolog�a sistem�tica, la siguiente entrada
taxon�mica: sistema de
vida org�nica, Solaria; bi�sfera, Marciana; rei�no, Jakobia; genus et species,
Gillevinia straata.
________
It was found that Levinia, the name given
on December 20, 2006 for the genus of the active specimens found in biosphere Marciana by the Viking Mission's Labeled Release life
detection experiments, whose detailed diagnoses (characteres
naturales) were provided by Klein et al. 1976, was
already in use for genera in biosphere
Terrestria. In Trilobites,
Levinia stands in fair use (Jell and
Adrain 2003) since Rusconi (1950) thus nomenclated a trilobite genus or
subgenus of indeterminate family from Mendoza, Argentina. Among
"coccidia", or protozoan parasites of the suborder Eimeriorina in
phylum Apicomplexa, the generic name Levinia
have been already proposed for the Isospora
species that utilize paratenic hosts species (cf. Dubey, 1977). Among Enterobacteriaceae,
Levinia
is also a mispelling/synonym of a later heterotypic
genus (Levinea Young
et al. 1971) synonym for
Citrobacter
Werkman and Gillen 1932 (cf. Farmer 1981, Brenner et al 1993).
Therefore, in order to ensure that generic names are unique across all
biological major taxa, the genus' name Levinia
is replaced and the genus that it formerly characterized in Marciana is hereby nomenclated
Gillevinia, novum genus. As a
consequence, the active
agent characterized by its reported behavior as responsible of the results of
the 1976 Viking Mission's Labeled Release experiment from now on formally
features, for systematic biology, the following taxonomical entry: organic life
system, Solaria; biosphere, Marciana; kingdom, Jakobia; genus et species, Gillevinia straata.
Referencias
Brenner,
D. J., Grimont, P. A. D., Steigerwalt, A. G., Fanning, G. R., Ageron, E. and
Riddle, C. F. (1993), "Classification of citrobacteria by DNA hybridization:
designation of Citrobacter farmeri sp.
nov., Citrobacter youngae sp. nov., Citrobacter braakii sp. nov., Citrobacter werkmanii sp. nov., Citrobacter sedlakii sp. nov., and three
unnamed Citrobacter genomospecies",
Int. J. Syst. Bacteriol. 43, 645-658
Dubey, J.
P. (1977), "Toxoplasma,
Hammondia, Besnoitia, Sarcocystis, and other tissue cyst-forming coccidia of
man and animals". In J. P. Krier (ed. ), Parasitic protozoa, vol. 3, p. 101�237.
Academic Press, Inc., New York,
Farmer
III, J. J. (1981), "The genus Citrobacter".
In: M. P. Starr et al. (eds), The procaryotes, a handbook on habitats, isolation,
and identification of bacteria. Springer-Verlag, Berlin, p. 1140-1147
Gillespie,
S. H. and P. M. Hawkey, eds. (2006), Principles and Practice of Clinical Bacteriology
(Second Edition). John Wiley & Sons, Ltd.
Klein,
H. P., N. H. Horowitz, G. V. Levin, V. I. Oyama, J. Lederberg, A. Rich, J. S.
Hubbard, G. L. Hobby, P. A. Straat, B. J. Berdahl, G. C. Carle, F. S. Brown,
and R. D. Johnson (1976), "The Viking Biological Investigation:
Preliminary Results", Science 194 (4260), 99-105. DOI:
10.1126/science.194.4260.99
Rusconi,
C. (1950), "Trilobitas y otros organismos del C�mbrico de Canota",
Revista del Museo de Historia Natural de Mendoza 4: 71-84.
Young, V.
M., Kenton, D. M., Hobbs, B. J. and Moody, M. R. (1971), "Levinea, a new genus of the family
Enterobacteriaceae", International Journal of Systematic Bacteriology 21:
58-63.
__________
Copyright � April 2007 Electroneurobiolog�a. Este
texto es un art�culo de acceso p�blico; su copia exacta y redistribuci�n por
cualquier medio est�n permitidas bajo la condici�n de conservar esta noticia y
la referencia completa a su publicaci�n incluyendo la URL (ver primera p�gina).
/ This is an Open Access article: verbatim copying and redistribution of this
article are permitted in all media for any purpose, provided this notice is
preserved along with the article's full citation and URL (first page).
revista
Electroneurobiolog�a
ISSN: ONLINE 1850-1826 - PRINT 0328-0446









